Un laboratoire spécial a nommé ce virus. Méthodes de tests de laboratoire pour identifier les virus et caractéristiques de déchiffrement des résultats de la recherche. Microscopie optique
Une flambée sans précédent de l’épidémie mortelle du virus Ebola en Afrique de l’Ouest, qui menace de se propager au continent européen. Le SIDA, qui détruit des dizaines de millions de personnes, et d’autres maladies terribles jusqu’alors inconnues touchant les humains, les animaux et les plantes. Où nous tombent-ils sur la tête ? Quel rôle jouent dans tout cela les laboratoires secrets de la CIA et des départements militaires américains ?
"C'est impossible ! Le cancer n'est pas contagieux ! Ce sont toutes des inventions, comme des « théories du complot » ou des rencontres avec des Martiens ! C’est ainsi que les autorités américaines ont répondu aux accusations du gouvernement vénézuélien selon lesquelles le grand leader de la révolution bolivarienne, Hugo Chavez, aurait été détruit en lui transmettant un virus du cancer.
Cependant, les experts estiment qu'un si grand nombre de dirigeants latino-américains (et de gauche !) tombés malades du cancer à peu près au même moment ne peut pas être expliqué par des causes naturelles. Parmi eux, aux côtés de Chavez, figurent le président argentin Néstor Kirchner, qui lui a succédé, Cristina Kirchner, le président brésilien I. Lula da Silva, qui est arrivé au pouvoir après lui, Dilma Rousseff, et le président paraguayen Fernando Lugo (renversé lors de la Seconde Guerre mondiale). un coup d'État de droite en 2012). , organisé par la CIA ; et diagnostiqué peu après avec un cancer du système immunitaire). Le dirigeant cubain Fidel Castro a survécu de justesse à un mystérieux cancer de l'intestin qui l'a frappé après le Sommet des peuples de 2006 dans la ville argentine de Cordoba.
Peu de gens savent que bien avant les expériences brutales des camps de concentration allemands pendant la Seconde Guerre mondiale, les Américains ont mené des expériences similaires sur les habitants de l’Amérique latine sous les auspices de l’Institut Rockefeller pour la recherche médicale.
L'un des fanatiques, Cornelius Rhodes, écrivait à son ami en 1931 : « Tout est merveilleux ici à Porto Rico, à l'exception des Portoricains. Ce sont sans aucun doute les dégénérés les plus sales et les plus paresseux de la race voleuse qui habite cet hémisphère. Pour la santé publique, certains moyens sont nécessaires pour tous les détruire. Et j'ai tout fait pour accélérer ce processus - j'en ai tué huit au cours des expériences et j'en ai infecté beaucoup avec le cancer. Ici, il n'y a pas d'assurance maladie ni de prestations sociales. C'est ce que admirent les médecins qui sont libres de guérir jusqu'à la mort et de torturer leurs malheureux patients.»
Le « Docteur » a injecté par voie intraveineuse des substances biologiques cancérigènes et au moins 13 patients sont morts à la suite de ces expériences cruelles.
Dans les années 1950, Rhodes devient directeur des programmes de recherche sur les armes chimiques et biologiques au centre militaire de Fort Detrick dans le Maryland, sur les sites d'essais du désert de l'Utah et du canal de Panama, puis rejoint la Commission américaine de l'énergie, qui expose les Américains sans méfiance à des radiations radioactives. déterminer le niveau de « rayonnement sûr » et l’apparition de tumeurs malignes à la suite de ces expériences.
Après la mort de Rhodes, l'American Cancer Association a créé un prix en son nom. Cependant, en 2004, à la suite des révélations scandaleuses de ses expériences sauvages, le président de l'association, S. Horwitz, a annoncé que la plus haute distinction décernée aux oncologues américains ne serait plus associée au nom de Rhodes en raison du « caractère controversé ». de ses activités. »
Il y avait une douzaine de ces canailles de la science aux États-Unis, et ils ont testé presque toutes les infections qu'ils ont inventées, d'abord en Amérique latine (sans oublier les expériences sur leurs propres citoyens). Après la guerre, le champ s'est rétréci du fait que beaucoup ont commencé à se tourner vers l'URSS pour obtenir une aide médicale et scientifique. Mais après l’effondrement de l’Union soviétique, des perspectives véritablement illimitées se sont ouvertes à ces écorcheurs.
Obama a déjà été contraint à plusieurs reprises de s'excuser auprès des pays d'Amérique latine pour les expériences menées sur des personnes dans les années 40 et 50, qui ont conduit à la propagation de la syphilis et d'autres maladies sexuellement transmissibles, à une stérilité massive et à diverses épidémies. Cependant, de telles excuses (seulement après la publication de preuves irréfutables !) ne feront pas revivre les millions de morts et de victimes du bioterrorisme américain, ni ne conduiront à l’arrêt de telles « expériences » à l’avenir (selon le principe « si pas attrapé, pas de voleur »).
Depuis la fin des années 60, le développement et la création accélérés de diverses modifications du virus du cancer ont commencé. Le travail a été coordonné avec le National Cancer Institute, qui a officiellement développé des traitements pour la « maladie du siècle » et a participé officieusement aux projets de la CIA visant à utiliser le virus du cancer à des fins militaires et politiques.
Malgré la cérémonie de signature en 1972 à Moscou, Londres et Washington de la Convention sur l'interdiction de la mise au point, de la fabrication et du stockage des armes bactériologiques (biologiques) ou à toxines et sur leur destruction (BTWC), les travaux à Fort Detrick battaient leur plein et en 1977, 60 000 litres de virus cancérigènes et immunosuppresseurs avaient été produits.
Les professeurs R. Purcell, M. Hillerman, S. Kragman et R. McCollum ont participé activement aux travaux, qui ont utilisé un « cocktail » du virus de l'hépatite B en combinaison avec une substance oncogène pour des expériences non seulement sur des macaques rhésus et des chimpanzés, mais aussi également contre les étudiants américains de la Willowbrook State School for Mentally Retarded Children.
En 1971, la société pharmaceutique américaine Lytton Bionetics a conclu des contrats avec plusieurs pays africains pour étudier des patients atteints de cancer atteints du lymphome de Birkett, associé à l'oncovirus infectieux d'Epstein-Barr, ainsi que de leucémie et de sarcome. Il est curieux que le lymphome de Birket ait été découvert pour la première fois dans l'ouest de l'Ouganda après que les laboratoires du National Cancer Center des États-Unis, ainsi que d'autres institutions médicales parrainées par Rockefeller, y aient travaillé.
L’un des experts, R. King, a déclaré dans les années 80 que des spécialistes américains avaient infecté des personnes atteintes de sarcome afin « d’isoler le génome du virus par récupération, hybridation, recombinaison de virus, mutations et autres techniques techniques ».
Lors des audiences du Comité sénatorial Church en 1975, le Dr Charles Senseney, qui travaillait au laboratoire de Fort Detrick, a admis que la CIA utilisait des substances biologiquement actives qui provoquaient des maladies cardiaques passagères et des cancers pour détruire des personnages indésirables. Il a montré des échantillons d'armes avec lesquelles les victimes visées avaient été infectées. Parmi eux se trouvaient un parapluie qui tirait des fléchettes miniatures lorsqu'il était ouvert, ainsi qu'une sarbacane spéciale pour tirer des aiguilles fabriquées à partir d'une substance toxique gelée. Aussi épaisses qu'un cheveu humain et longues de plusieurs millimètres, ces aiguilles traversaient le tissu des vêtements sans dommage et, une fois injectées, provoquaient une sensation douloureuse pas pire qu'une piqûre de moustique, se dissolvant instantanément sous la peau.
Parmi les « nouveaux produits » des bioterroristes américains ont également été présentés des aérosols destinés à infecter des « cibles » avec des maladies mortelles après pulvérisation depuis des avions, ainsi que des « virus sauteurs » propagés par des insectes (puces, araignées, moustiques) qui sautent ou s'envolent d'animaux infectés. aux humains. La CIA est également devenue « pionnière » dans les méthodes d’infection : par injections, inhalations, contact avec la peau de vêtements contaminés, par le système digestif en mangeant, en buvant et même en utilisant du dentifrice.
Un certain nombre d'experts estiment que l'un des premiers dirigeants politiques détestés par les États-Unis à avoir été infecté par une nouvelle arme biologique contre le cancer a été le président de l'Angola, Agostinho Neto. Il est décédé à l'hôpital clinique central de Moscou en 1979, à l'âge de 57 ans, d'une forme jusqu'alors inconnue de cancer fulminant. Une autre victime a été l'ancien président du Chili, Eduardo Frey, qui s'est ouvertement opposé au protégé américain, le général Pinochet. Frey est décédé dans un hôpital de Santiago en janvier 1982, contractant une maladie fulminante inconnue après avoir subi un examen médical de routine.
Ainsi, peut-être que dans 50 ans, les archives de la CIA seront déclassifiées et que les secrets de la mort d'Hugo Chavez et d'autres dirigeants mondiaux seront connus. Il existe une telle quantité de documentation sur l’utilisation de virus cancéreux par les agences de renseignement américaines que l’existence de ces armes ne soulève aucune question. La seule question est de savoir comment cela a été « introduit » et qui en était l’auteur direct.
* * *
« Dans les 5 à 10 prochaines années, il sera possible de créer un virus synthétique qui n'existe pas du tout dans la nature et qui ne peut pas être supprimé par le système immunitaire humain ; Les nouveaux virus créés artificiellement seront inaccessibles aux médicaments ; il est inutile d’utiliser les moyens conventionnels de traitement des maladies infectieuses, les antibiotiques, les vaccins et les antidotes contre celles-ci. » Une déclaration aussi sensationnelle a été faite par le virologue en chef de l'armée, D. MacArthur, s'exprimant en 1969 devant les commissions du Congrès américain («Commission Sykes»), censées faire des recommandations sur l'allocation des fonds budgétaires à l'armée. Et il a demandé peu – seulement environ 10 millions de dollars !De l'argent a été alloué et des centaines de chercheurs et d'experts ont été impliqués dans les travaux. L’un des créateurs du virus du SIDA était apparemment le Dr Robert Gallo, qui a même reçu en 1987 un brevet du ministère américain de la Santé établissant sa priorité dans l’invention d’un « virus qui supprime le système immunitaire humain ».
La maladie a échappé aux laboratoires et a été découverte pour la première fois au printemps 1981 en Californie (USA). Et cela n’a rien à voir (comme les Américains tentent de nous le convaincre) avec l’Afrique et les « petits singes verts ».
En mai 1987, un article paru dans le London Times affirmait que la vaccination contre la variole en Afrique (initiée par les « humanistes » du ministère américain de la Santé) avait provoqué une épidémie de SIDA. Et des millions de personnes ont été vaccinées ! Ensuite, une « vaccination » similaire a été réalisée en Haïti, au Brésil et dans d’autres pays.
Les accusations portées contre les États-Unis pour avoir fabriqué le virus du SIDA ont commencé au milieu des années 80. Le professeur à l'université Humboldt de Berlin, Jakob Segal, a affirmé que le virus est "le produit d'une expérience menée en laboratoire dans le but de créer une arme biologique". Dans les médias américains, tout cela a été présenté comme de la « propagande soviétique ». Mais dans les années 90, le Dr Gallo lui-même a annoncé qu'il avait testé une autre souche « alternative » du SIDA, qui peut pénétrer dans l'organisme par les cellules épithéliales (c'est-à-dire par la peau), augmentant ainsi le risque de contracter la maladie en pulvérisant le produit. substance active dans l'atmosphère.
Le Dr S. Monteith a été l’un des premiers à décrire, dès 1981, l’énorme potentiel épidémique du nouveau virus, les conséquences potentiellement catastrophiques de son utilisation par « l’élite mondiale », et à prouver également son caractère artificiel.
Et cette nouvelle qualité a jusqu’à présent empêché toute tentative de création d’un vaccin contre le SIDA. C'est pourquoi, au fil des années, aucun médicament efficace n'a été créé contre cette maladie.
Le nombre de personnes infectées par le SIDA est encore inconnu, car même aux États-Unis, le gouvernement empêche toute initiative visant à un décompte, même approximatif. Selon diverses estimations, entre 50 et 100 millions de personnes seraient infectées par le SIDA. Surtout en Afrique - dans certains pays (Ouganda, Kenya), plus de 50 % de la population souffre de cette terrible maladie.
On estime qu’environ 40 millions de personnes sont mortes du SIDA à ce jour – presque le même nombre que pendant la Seconde Guerre mondiale !
* * *
Selon l'Organisation mondiale de la santé, plus de 600 personnes infectées par Ebola sont déjà mortes à l'ouest du « continent noir ».L’épidémie actuelle de la maladie est devenue la plus importante de l’histoire des observations médicales.
Au Nigeria, au Libéria et dans d'autres pays africains, des cordons spéciaux sont installés aux frontières et des médecins surveillent attentivement toutes les personnes entrant et sortant. La fièvre Ebola est considérée comme une maladie mortelle à laquelle les humains, les primates et les porcs sont les plus sensibles. Il n’existe pas de vaccin contre cela.
L'épidémie a débuté en Guinée en mars de cette année. À ce jour, la maladie se propage à de nouveaux territoires en Sierra Leone, au Libéria et au Mali. On craint qu’elle ne se propage non seulement dans toute l’Afrique de l’Ouest, mais qu’elle pénètre également en Europe.
Il est curieux que dans les foyers de l'épidémie, les cas d'attaques de riverains contre les locaux de l'organisation internationale Médecins sans frontières se soient fortement multipliés. Les habitants accusent les médecins d'avoir introduit le virus dans la région. Des manifestations massives ont eu lieu contre les gouvernements africains qui ne font rien pour corriger la situation.
Les pogroms des bureaux d’une « organisation internationale respectée » sont présentés dans la presse occidentale comme des exemples « d’irrationalité et d’absurdité ». Par ailleurs, « Médecins sans frontières » vante de toutes les manières possibles ses principes éthiques, assurant être « toujours proches des victimes ». Mais ne s’agit-il pas de leurs propres victimes, comme le croient les Africains « déraisonnables » ?
Pourquoi les médecins occidentaux ne quittent-ils pas obstinément la Guinée, le Libéria, le Mali et la Sierra Leone ? Après tout, ces pays sont plongés dans le chaos des guerres civiles et des conflits, auxquels les pays européens et les États-Unis participent activement. La France à elle seule a dépensé des centaines de millions d’euros pour des opérations militaires au Mali.
Tout – pour restaurer le pouvoir colonial en Afrique de l’Ouest et du Nord. Et ce sont ces territoires qui sont « débarrassés » de la population locale lors des épidémies d'Ebola et d'autres maladies infectieuses. De plus, étonnamment, seuls les résidents locaux souffrent, mais pas les « casques bleus » français.
Et « médecins sans frontières » ne transfère pas de médicaments et de matériel aux autorités locales et ne quitte pas la zone de conflit. C’est précisément ce qui donne aux résidents locaux de bonnes raisons de soupçonner les « esculapiens » étrangers d’être ceux qui propagent de nouvelles souches d’infection parmi les Africains.
Selon de nombreux experts, de nouvelles armes « ethniques » y sont testées, qui agissent de manière sélective - uniquement sur les Africains. Mais il semble qu'il y ait des modifications pour d'autres groupes raciaux et ethniques. En 2006, l'un des principaux virologues américains, Eric Pianka, s'exprimant lors d'une cérémonie à l'Université du Texas, a déclaré qu'avec l'aide d'une nouvelle souche de la fièvre Ebola (selon ses mots, « avec une létalité fantastique »), il est possible « pour le bien de la planète » pour réduire l’humanité de 90 %. Les virologues américains présents dans la salle se sont unanimement levés et lui ont fait une standing ovation...
* * *
Depuis les années 70, un développement accéléré des « armes ethniques » a été réalisé aux États-Unis. Et comme le pensent de nombreux experts, de nouvelles souches de virus mortels ont été inventées et ne peuvent se propager que dans un certain environnement ethnique.Ainsi, le « SRAS » touche surtout les Chinois et les résidents d'Asie du Sud-Est, Ebola et le SIDA - les Africains. Les scientifiques israéliens tentent de créer une arme biologique similaire dirigée contre les Arabes.
La British Medical Association a récemment déclaré que « les progrès progressifs de la génétique pourraient conduire à un nettoyage ethnique d’une ampleur sans précédent dans les années à venir ».
L’idée d’établir une « domination biologique sur le monde » ne mûrit plus seulement dans l’esprit des virologues cannibales fous, mais dans les calculs des politiciens, des stratèges militaires et des experts ! Ainsi, cette idée a été récemment exprimée par de respectables politiciens néoconservateurs américains dans le rapport « Nouvelles frontières pour la défense américaine ».
Il affirme que, bien entendu, la domination militaire sur le monde doit avant tout être assurée par des missiles balistiques et de croisière, des avions et sous-marins radiocommandés (« drones ») et des armes satellitaires. Mais parallèlement, « dans les années à venir, l'art de la guerre dans les airs, sur terre et sur mer sera complètement différent de l'art actuel, et les batailles se dérouleront dans de nouvelles dimensions - dans l'espace, le cyberespace, ainsi que comme au niveau intracellulaire et microbien. Et en outre, il est dit que « des formes avancées d’armes biologiques, qui sélectionneront certains génotypes humains comme cibles, seront capables de faire passer cette direction du monde de la terreur à la place qui lui revient parmi les moyens politiquement justifiables » !
* * *
Les autorités américaines ont bien tiré les leçons du projet Manhattan, en particulier du transfert de données sur les armes atomiques vers l'Union soviétique par les plus grands physiciens du monde. Les scientifiques américains ne l'ont pas fait pour de l'argent, mais sur la base d'une évaluation sobre de leur gouvernement, qui n'hésiterait pas à bombarder l'URSS et tous les autres concurrents potentiels sur la voie de la domination mondiale.C’est pourquoi les développeurs de nouveaux virus sont désormais soumis aux règles les plus strictes pour éliminer les « témoins indésirables ». Le taux de mortalité parmi eux est des dizaines de fois supérieur à la moyenne statistique.
Des experts américains indépendants ont dénombré plus d'une centaine de décès « mystérieux » (dans des accidents d'avion et de voiture, dus à des maladies « inconnues », des « accidents ») parmi les virologues et microbiologistes travaillant sous contrat pour la CIA et le ministère de la Défense.
En 2001, immédiatement après l’explosion des « tours à crêpes », tous les Américains ont été alarmés par la nouvelle de lettres contenant des spores d’anthrax envoyées aux rédactions de magazines, de journaux, de chaînes de télévision et de personnalités politiques. 17 personnes ont été infectées, cinq sont mortes. Ces lettres ont été la principale raison du tournant politique qui a dirigé l’agression américaine contre l’Irak. Al-Qaïda est tombé dans l’oubli et tous les médias ont rapporté que « la plus grande attaque biologique de l’histoire des États-Unis » avait été organisée par Saddam Hussein.
Lorsque cette torsion a été cimentée (et utilisée plus tard pour accuser Hussein de développer des armes biologiques, ce qui est devenu l'un des arguments en faveur de l'invasion de l'Irak), il est rapidement devenu clair que la souche du virus ne pouvait être obtenue qu'au laboratoire de la CIA à Fort. Détruire. Là, ils ont trouvé un «maillon faible» - le virologue Bruce Ivins, qui, étant un fervent catholique, se plaignait souvent de ne pas aimer son travail pour des raisons religieuses. Et en juillet 2008, il se serait suicidé en avalant des drogues puissantes. Après cela, le FBI l'a qualifié de « terroriste fou » qui envoyait des lettres infectées. Aucune autopsie n’a été pratiquée, aucune enquête n’a été menée et l’affaire a été rapidement classée.
Il est curieux qu'il ait répété le sort de l'un des principaux microbiologistes des années 50, Frank Olson, qui a également travaillé sur l'anthrax et a présenté sa démission de Fort Detrick, ne voulant pas participer au développement d'armes mortelles. Et quelques jours plus tard, en novembre 1953, selon le rapport du FBI, « en état de dépression nerveuse, il sauta du 10e étage de l'hôtel Pennsylvania ».
L’un des cas les plus célèbres est celui du « suicide » du plus grand expert britannique en armes biologiques, David Kelly. Il s'est rendu en Irak des dizaines de fois dans le cadre de diverses missions d'inspection de l'ONU. Après l'invasion, il a fait une (première !) déclaration sensationnelle selon laquelle tous les « documents » sur la présence des armes chimiques et bactériologiques de S. Hussein, présentés par les autorités américaines et britanniques à l'ONU et qui ont servi de prétexte à la guerre , étaient des « contrefaçons grossières ». Il a été convoqué au Parlement, où lors des auditions, il n'a pratiquement pas été autorisé à ouvrir la bouche, l'attaquant de reproches et d'accusations.
Quelques jours plus tard, le 17 juillet 2003, il partit, comme toujours, faire une promenade matinale et son corps fut découvert le lendemain à un kilomètre et demi de chez lui. Le rapport officiel indique qu'il s'est suicidé en avalant 30 somnifères, puis en se coupant une veine du poignet gauche avec un couteau. Mais les médecins ambulanciers (ne connaissant apparemment pas cet « ordre ») ont constaté qu’il n’y avait pas de sang sous le cadavre. En conséquence, Kelly s'est empoisonné, s'est coupé une veine, puis, saignant, il est arrivé lui-même à l'endroit où il a été retrouvé !
Aux États-Unis, l'un des événements les plus notoires a été l'accident d'avion de mars 2002, au cours duquel Stephen Mostow, un éminent virologue qui travaillait au Colorado Medical Center, est décédé. On l'appelait « Monsieur Grippe » car il était principalement spécialisé dans cette maladie.
Parmi les morts se trouvaient de nombreuses personnes de notre pays qui, pour diverses raisons, sont allées « chercher le bonheur » en Occident. Le plus marquant a été une « crise cardiaque » en 2001 chez le microbiologiste V. Pasechnik, qui était en bonne santé. L’Occident l’a utilisé (comme beaucoup d’autres Russes) à 200 % – à la fois comme spécialiste et comme « dénonciateur des terribles conspirations du Kremlin contre les États-Unis et le monde libre tout entier ».
En 1989, il part en Angleterre et y travaille dans l'un des centres de virologie. En cours de route, il a gagné de l'argent en racontant des histoires sur « l'arme biologique binaire » soviétique appelée « Novitchok », selon laquelle tous les virus connus étaient maîtrisés depuis longtemps dans les laboratoires secrets du KGB et de nouveaux étaient déjà apparus. Ils peuvent provoquer des « maladies monstrueuses » telles que la sclérose en plaques et l’arthrite chez les Américains sans méfiance.
Ces histoires d’horreur étaient utiles car elles fournissaient une excuse pour retirer des fonds budgétaires pour la « biodéfense » (en réalité, pour le développement de nouvelles souches mortelles). Mais ensuite ils ont décidé que le bavard Pasechnik parlait trop du centre de virologie de Sailsbury, où il a travaillé pendant 10 ans, et l'ont envoyé dans un autre monde...
* * *
« La fusée de Poutine », « la main de Moscou », « Poutine, tu as tué mon fils ! - Les magazines et journaux occidentaux ont fait la une de ces gros titres après qu'un avion de ligne Boeing reliant les Pays-Bas à Melbourne a été abattu dans le ciel ukrainien le 17 juillet de cette année. Cette hystérie a commencé immédiatement après le discours du président américain Obama, qui a déclaré qu'il s'agissait d'un « crime aux proportions inimaginables » et en a imputé la responsabilité à la Russie. Immédiatement entre les mains des secrétaires de presse de la Maison Blanche et du Département d'État, sont apparues des photographies floues reçues de la CIA et « indiquaient de manière irréfutable » que l'avion de ligne avait été abattu par un missile russe Buk.Cet événement a été à l'origine du déploiement à grande échelle de sanctions économiques contre la Russie, de l'implication des pays de l'UE (avant le désastre, ils hésitaient à soutenir les États-Unis), de l'utilisation de presque tous les moyens de guerre interdits pour réprimer la résistance. à Novorossiya (bombes au phosphore, missiles balistiques, ogives à fragmentation, etc.), la mise en œuvre de plans visant à constituer un bloc militaire anti-russe avec la participation de l'Ukraine, de la Moldavie, de la Pologne, de la Géorgie et des pays baltes.
Seulement un mois plus tard, des informations ont commencé à apparaître selon lesquelles des trous dans le cockpit et le fuselage prouvaient que l'avion avait été abattu en l'air, très probablement par un chasseur de l'armée de l'air ukrainienne. Cette version est confirmée par un brusque changement de route du Boeing juste avant la catastrophe. Cependant, l’affaire est déjà accomplie, tous les médias occidentaux ont immédiatement oublié l’avion, et les sanctions et une guerre à grande échelle contre le peuple russe dans l’est de l’Ukraine non seulement sont en vigueur, mais continuent de s’intensifier.
Il y a tous les signes d'un « événement déclencheur » ou d'un « incident falsifié » (incident sous fausse bannière) - c'est ainsi que les maîtres des provocations de la CIA appellent des attaques terroristes visant à orienter l'opinion publique dans la direction nécessaire aux États-Unis. Les États, pour déclencher une chaîne d'événements qui mèneront à la réalisation des objectifs de « l'empire ». Cela a toujours été le cas dans l'histoire des États-Unis : l'explosion de son propre cuirassé Maine, qui est devenue le prétexte pour déclarer la guerre à l'Espagne en 1898 ; le naufrage prévu du paquebot Lusitania pour entrer dans un moment avantageux de la Première Guerre mondiale ; suppression délibérée des informations sur l'attaque japonaise imminente contre la base américaine de Pearl Harbor en 1941 pour entrer dans la Seconde Guerre mondiale ; provocation avec le bombardement du destroyer américain Maddox dans le golfe du Tonkin pour déclarer la guerre au Vietnam en 1964 ; le bombardement des Twin Towers en 2001 pour déclencher la « guerre contre le terrorisme » et préparer l’invasion de l’Irak et de l’Afghanistan.
Comme c’est souvent le cas lors de telles attaques terroristes, ce n’est pas un seul, mais plusieurs objectifs qui sont poursuivis. Dans ce cas, l'information selon laquelle il y avait plus d'une centaine de microbiologistes à bord du MH17 qui se rendaient au congrès international sur le sida en Australie est d'un grand intérêt. Et parmi eux se trouve J. Lange, un virologue de premier plan à l’Université d’Amsterdam.
« La perte irréparable du plus grand visionnaire et titan de l’étude du sida », « la mort tragique du plus grand expert mondial dans le traitement de la maladie du siècle », ont été écrites dans des nécrologies publiées dans des revues scientifiques. Et en effet, le laboratoire de Lange a pris une position de leader dans l’étude du SIDA et des méthodes de traitement, y compris l’utilisation combinée de médicaments et de thérapies antirétrovirales, et a développé des moyens de prévenir la transmission du virus de la mère à l’enfant. Pendant plusieurs années (2002-2004), il a dirigé l'organisation internationale de lutte contre le sida. À ses côtés se trouvaient ses collègues néerlandaises Jacqueline van Tongeren, M. Adriana de Schutter, L. Vann Mens et d'autres scientifiques. Il est possible qu'ils aient apporté avec eux les résultats de nombreuses années de travail, peut-être même un remède tant attendu contre cette maladie monstrueuse - après tout, peu de temps avant la conférence, les employés de Lange ont déclaré que son discours devrait faire sensation dans le monde scientifique. .
Dans le même Boeing (par hasard, semble-t-il), se trouvait le représentant de l'Organisation Mondiale de la Santé (OMS), Glenn Thomas, qui a été « condamné à une amende » pour avoir accordé une interview dans laquelle il évoquait le rôle criminel de son organisation dans la propagation du virus. l'épidémie d'Ebola en Afrique de l'Ouest.
En détruisant les chercheurs européens sur le SIDA, ainsi qu'un honnête fonctionnaire de l'OMS, les Américains ont ainsi donné une leçon à tous ceux qui s'efforcent sincèrement de guérir le SIDA et Ebola : « Il n'est pas nécessaire de traiter et de prévenir ces maladies, elles sont très utiles pour nous pour la destruction de la populace humaine qui prolifère.
Ce n'est pas un hasard si de nombreux articles rappellent qu'en 1998, un avion de Swissair s'est écrasé au-dessus de l'Atlantique, transportant l'un des brillants chercheurs sur le sida, Jonathan Mann, et son épouse M. L. Clements, également célèbre virologue. Mann dirigeait la structure de l'OMS chargée de lutter contre le sida et, comme l'ont écrit ses collègues, sa mort a porté un coup dur à tous les plans visant à organiser la lutte contre cette terrible maladie. Les causes de la catastrophe n’ont pas encore été élucidées (aucun des experts sérieux ne croit à la version officielle selon laquelle un mégot de cigarette des pilotes est tombé, ce qui a provoqué un incendie à l’intérieur de l’avion).
* * *
Les États-Unis utilisent contre nous un énorme arsenal d'armes biologiques : les OGM et les plantes et organismes transgéniques (dont beaucoup, selon les experts occidentaux, provoquent la suppression du système immunitaire, le cancer, l'infertilité et les maladies cérébrales), organisent chaque année des dizaines d'épidémies de nouveaux Les virus de la grippe, les maladies animales (« porcine » et « grippe aviaire »), les plantes, propagent diverses maladies allergiques, vendent des médicaments et des vaccins aux « effets secondaires » qui nous sont inconnus, des additifs alimentaires, etc. De plus en plus de nouveaux virus se développent : le « hantavirus » mortel, le « virus tueur australien » recombinant basé sur la variole, une nouvelle génération de maladies « non mortelles » (seulement complètement « invalidantes »), des « biorégulateurs » capables de créer une dépression à grande échelle, en modifiant le rythme cardiaque , et conduisant à l’insomnie. Il est possible que des « signets » biologiques soient créés – des virus latents qui devraient être activés après un certain temps.Des laboratoires biologiques militaires américains se créent autour de la Russie : en Géorgie (où, selon les experts, l'épidémie de peste porcine est née en 2013), au Kazakhstan, au Kirghizistan et dans les États baltes. Les autorités américaines consacrent d’énormes sommes d’argent au développement de nouveaux virus et à la biodéfense (plus de 6 milliards de dollars sont dépensés chaque année pour le seul programme Bioshield).
Dans notre pays, après l'effondrement de l'Union soviétique, pendant longtemps, presque aucune attention n'a été accordée à ce domaine le plus important de la protection du pays. Instituts et centres fermés, jeunes spécialistes partis vers l'Occident. Il ne reste que des passionnés et des scientifiques âgés qui travaillent pour de maigres salaires (18 000 chercheurs seniors, 27 000 professeurs, docteurs en sciences).
Bâtiments vétustes, équipements vétustes, « pression supplémentaire » de la part des responsables libéraux. C’est arrivé au point qu’en 2000, pour « sous-paiement », Mosenergo de Chubais a tenté de couper l’électricité à l’Institut de virologie Ivanovsky. Non seulement une collection unique de micro-organismes serait détruite, mais certains échantillons de virus pourraient s’échapper dans l’atmosphère ! Ensuite, ce n’est que par miracle que nous avons réussi à combattre les « managers efficaces ». Et le coup final a été porté par la « réforme » de l’Académie des sciences de Russie – en fait, sa liquidation et le transfert de la direction entre les mains d’un comptable « efficace » de Krasnoïarsk.
Personne n'a interféré avec la véritable chasse aux agents de la CIA contre les scientifiques patriotes, qui ont simplement été détruits sur le territoire de notre propre pays ! En janvier 2002, A. Brushlinsky, membre correspondant de l'Académie des sciences de Russie, directeur de l'Institut de psychologie, psychologue et biologiste, auteur d'ouvrages sur la reconnaissance des terroristes, a été battu à mort avec des battes de baseball (afin qu'ils sachent où se trouvait l'ordre). pour liquidation !) et étranglé devant l'entrée de sa maison à Moscou. Deux ans après sa mort, son adjoint, le professeur V. Druzhinin, a été tué.
En novembre 2002, le professeur B. Svyatsky, spécialiste des infections infantiles de l'Université médicale d'État de Russie. Pirogov. Membre correspondant de l'Académie russe des sciences médicales, virologue et microbiologiste de premier plan, spécialiste des armes biologiques L. Strachunsky, a été battu à mort avec des battes de baseball en 2005 dans sa chambre de l'hôtel Slavyanka de Moscou. En 2006, le généticien et biologiste, membre correspondant de l'Académie russe des sciences L. Korochkin a été tué.
Une perte énorme pour la microbiologie nationale a été la mort du chef du département de microbiologie de l'Université médicale d'État de Russie, le professeur V. Korshunov, l'un des plus grands virologues au monde, spécialiste reconnu des « anti-armes » biologiques. Le scientifique de 56 ans a été battu à mort par des « hooligans inconnus » en 2002, quelques jours après la publication d'un article de journal affirmant que le scientifique était sur le point de faire la plus grande découverte : un vaccin universel contre toute arme biologique ! À la suite de la mort de Korshunov, les travaux dans le domaine scientifique le plus important ont été arrêtés. En Russie, des centaines, voire des milliers de personnes ont été condamnées à la mort à cause de l’arrêt des recherches.
Les pages tragiques de l’histoire moderne nous convainquent que les États-Unis sont capables de toutes les actions les plus barbares et criminelles dans leur désir maniaque de domination mondiale. Il est significatif que les pays qu'ils envahissent sous prétexte de « protection des droits de l'homme », de « l'humanisme » et de la « démocratie » deviennent non seulement le théâtre des guerres civiles les plus aiguës, mais sont également accompagnés d'épidémies de diverses nouvelles, auparavant maladies inconnues. Au Vietnam, en Yougoslavie et en Irak, d'énormes masses de personnes ont été exposées à des substances mutagènes, ce qui a entraîné de terribles conséquences. De terribles malformations chez les bébés, la création de toute une génération de dégénérés, des changements irréversibles au niveau génétique qui affecteront toutes les générations futures - telles sont quelques-unes des conséquences des « actions humanitaires ».
De plus, les organisations internationales, dont l’ONU, actuellement entièrement sous contrôle américain, jouent le rôle de « couverture » dans la mise en œuvre de ce génocide. L’Organisation mondiale de la santé (OMS), Médecins sans frontières et d’autres organismes auparavant faisant autorité rédigent leurs « rapports objectifs » sous la dictée de l’Occident, et on ne peut plus leur faire confiance. Ils ont agi en collaboration avec les agresseurs en Irak, en Afghanistan et en Libye.
À la veille de l’invasion américaine de l’Irak, ils ont docilement conclu que Saddam Hussein possédait « d’énormes stocks d’armes biologiques et chimiques », ce qui a constitué l’un des principaux arguments pour inciter les États-Unis à déclencher une guerre. L'année dernière, ils ont accusé le gouvernement syrien d'avoir utilisé des armes chimiques et biologiques contre sa population lorsqu'environ 300 personnes ont été tuées en août par du gaz neurotoxique sarin dans une banlieue de Damas. Bien qu'à cette époque, des preuves solides aient été obtenues selon lesquelles le sarin était utilisé par des militants d'Al-Qaïda, elles n'avaient pas été obtenues nulle part, mais dans des entrepôts américains.
* * *
La destruction impitoyable des concurrents et, en fait, la tyrannie biologique des États-Unis détruisent la souveraineté des pays périphériques du monde, les obligeant à compter sur l’aide, l’expertise et les médicaments de l’étranger. Une telle dépendance coloniale porte atteinte à la sécurité des peuples, en fait des otages de l’Occident, des « rats de laboratoire » destinés à diverses expériences médicales et biologiques dirigées contre leur santé et leur vie.Le seul contrepoids à l’empire bioterroriste peut être le rejet du « mondialisme » vicieux et la construction d’un monde multipolaire. Tous les pays doivent, étape par étape, refuser de coopérer avec les États-Unis et l’OTAN, les organisations internationales pro-américaines existantes. Il est nécessaire de conclure des accords au niveau interétatique. Ainsi, en Afrique, les États doivent travailler ensemble pour lutter contre les nouvelles souches introduites d’Ebola. En Asie du Sud-Est - contre le nouveau syndrome le plus aigu du « SRAS ». C’est au niveau national que nous devons prendre soin de notre science, créer nos propres instituts et laboratoires nationaux, de puissants centres scientifiques pour contrer les armes virales et génétiques.
Nikolaï Ivanov
Étapes du diagnostic en laboratoire des maladies virales. Création d'un laboratoire de virologie.
1) Indication (détection) d'un virus dans du matériel pathologique :
Méthodes expresses :
a) Détection des virions :
(1) Microscopie électronique ;
(2) Microscopie optique (variole) ;
b) Détection des Ag viraux dans les réactions sérologiques (RIF, ELISA, RSK, RDP, RNGA) ;
c) Détection des corps d'inclusion par microscopie optique et à fluorescence ;
d) Détection des acides nucléiques viraux à l'aide de sondes PCR et ADN ;
e) Détection des hémagglutinines dans les RHA ;
f) Détection de l'activité infectieuse du virus dans un échantillon biologique.
2) Isolement (isolement) du virus du matériel pathogène. L'isolement est réalisé quels que soient les résultats de la première étape à l'aide d'un test biologique en trois passages « à l'aveugle ». Passage est l’infection d’un système vivant afin d’obtenir une nouvelle population du virus. Passage "aveugle"– infection sans signes visibles de reproduction virale. Après trois passages, le virus s'accumule dans les cellules d'un système vivant, ce qui s'accompagne de l'apparition de signes de reproduction, visibles au niveau du macroorganisme. Par exemple, quand test biologique sur les animaux: signes cliniques, décès, changements pathologiques ; sur embryons de poulet– décès, modifications pathologiques, hémagglutination ; sur culture de cellules– CPP, hémadsorption, plaques, RIF, etc. De tels systèmes vivants infectés sont définis comme un essai biologique positif. Cependant, il n’est pas encore possible de déterminer avec précision le type d’agent infectieux. Par conséquent, le matériel pathologique est sélectionné à partir d’un échantillon biologique positif, qui est classiquement considéré comme secondaire, c’est-à-dire sélectionné à partir d’un système vivant présentant des signes d’essai biologique positif. Une suspension contenant le virus ou des prélèvements d'empreintes digitales (virus isolé) en sont préparés.
3) Identification (détermination du type) du virus isolé dans des réactions sérologiques ou par analyse PCR. Dans de rares cas, une identification par d'autres caractères est possible, par exemple par des inclusions intracellulaires (corps de Babes-Negri pour la rage).
4) Si nécessaire, preuve du rôle étiologique du virus isolé. À cette fin, des réactions sérologiques sont utilisées, dans lesquelles le virus isolé est utilisé comme antigène et des échantillons appariés de sérum sanguin dans des dilutions en série au double sont utilisés comme anticorps. Un résultat positif, prouvant le rôle étiologique du virus isolé, est une augmentation du titre d'anticorps dans le deuxième échantillon de sérum sanguin de 4 fois ou plus par rapport au premier.
5) Diagnostic rétrospectif. À cette fin, des sérums sanguins appariés prélevés au stade de récupération sont utilisés, qui sont testés dans des réactions sérologiques avec un antigène spécifique standard conformément au diagnostic préliminaire d'une maladie virale. Une augmentation de 4 fois ou plus du titre d’anticorps dans le deuxième échantillon par rapport au premier indique un processus infectieux actif se produisant dans le corps de l’animal pendant la période de prélèvement sanguin. Dans ce cas, la maladie est causée par le virus contre lequel une augmentation du titre d'anticorps a été établie dans des sérums appariés.
Création d'un laboratoire de virologie.
Pour organiser un laboratoire de diagnostic, utilisez un compartiment isolé composé d'au moins 5 à 6 pièces.
Une salle lumineuse est réservée au laboratoire. Les locaux destinés à travailler avec du matériel viral doivent être bien éclairés et constitués d'un pré-box et d'un box, séparés par une cloison vitrée avec des portes. Seules les tables, chaises et accessoires de travail sont placés dans les cartons. La surface des tables est recouverte d'acier inoxydable, de plastique ou de verre, et des lampes bactéricides sont installées au-dessus du plan de travail. Un tapis désinfectant en éponge de caoutchouc imbibé d'une solution désinfectante est placé à l'entrée de la box. La salle de pré-boxe contient des vêtements et du matériel stériles adaptés aux besoins de la boxe. Le laboratoire est équipé d'eau froide et chaude et d'une ventilation avec apport d'air stérile.
Pour enregistrer le matériel pathologique entrant, il est prévu réception, où sont placés plusieurs tables recouvertes de tôles galvanisées et des récipients contenant des solutions désinfectantes (3% de chloramine, hydroxyde de sodium ou 5% de phénol).
Dans la salle de prétraitement des matériaux ( ouverture) ouvrez les cadavres et sélectionnez le matériel pour des recherches ultérieures.
Les débarras sont équipés en fonction de leur destination.
Dans un autoclave, les plats, les milieux de culture, les équipements et les milieux de culture sont stérilisés et le matériel infectieux est neutralisé. Il est nécessaire de disposer de deux autoclaves : pour les matériaux propres et pour ceux infectés.
La buanderie est conçue pour laver la vaisselle, le matériel et les appareils électroménagers.
Le vivarium doit disposer d'un service de quarantaine, de locaux pour animaux sains et expérimentaux et de locaux techniques.
Pour tout type de laboratoire de virologie, un élément obligatoire du laboratoire est une boîte de table ou, mieux encore, une boîte avec alimentation en air laminaire.

46. Cultures cellulaires et leurs types. Système dans lequel les cellules, tissus ou organes prélevés sur le corps conservent leur viabilité pendant au moins 24 heures. Survivre : dans lequel les cellules conservent uniquement leur activité vitale inhérente sans se reproduire. En croissance : conservent leur activité vitale inhérente et sont capables de se multiplier. Selon la nature de la croissance, ils sont divisés en 3 groupes : suspension ; plasma (cultures de morceaux de tissus fixés) ; une seule couche. Ceux à couche unique sont divisés en 4 groupes : primaire trypsinisé ; sous-cultures ; semi-sautable et entrelacable. Suspension: poussent sous forme de suspensions, les cellules se multiplient avec un milieu spécial et un mélange constant à l'aide de rouleaux. Les cellules se développent sur toute la surface du matelas. Un grand nombre de cellules pour les vaccins. Plasma: morceaux de tissus fixés par plasma, c'est la culture tissulaire. Il est obtenu en fixant un morceau de tissu sur un verre virologique, puis en y ajoutant du milieu de fosse et en le cultivant ; dans ce cas, la croissance cellulaire est enregistrée le long de la périphérie d’un morceau de tissu. Utilisé pour obtenir des morceaux de tissu. Une seule couche: Pour indiquer un virus. Il est obtenu à partir de tissus ou d'organes en les traitant avec de la trypsine. Les repiquages sont obtenus à partir des primaires par greffage. Ensuite, semi-transplanté par transplantations multiples. Ils possèdent un ensemble diploïde de chromosomes. Peut survivre diploïde en fonction de l'âge ou du tissu à partir duquel la culture cellulaire a été obtenue. Si l'embryon a jusqu'à 80 jours. Pour un adulte – pas plus de 25 greffes. Il n'y en a pas plus de 5. Les transplantées sont des cellules mutées et cancéreuses. Ils durent un nombre infini de fois. Ce sont des cellules transformées d'une tumeur cancéreuse. Hela est la culture cellulaire continue la plus connue depuis 1956. Cette culture est présente dans tous les laboratoires du monde. Il est adapté à de nombreux pathogènes. Les animaux premiers-nés présentent de nombreux avantages : ils ne meurent pas ; taux de croissance plus élevé; ils sont tous génétiquement homogènes. Chez les laboptoria, ils sont entretenus par réensemencement d'un récipient à l'autre.
59. DPC. Il s'agit d'une méthode permettant d'indiquer la présence du virus dans une culture cellulaire. Le CPD fait référence à tout changement dans les cellules d'une culture cellulaire sous l'influence d'un virus qui s'y reproduit. J'utilise un faible grossissement lorsque je regarde la couche supérieure du matelas. Comparez les cellules infectées avec celles non infectées. Les différences peuvent s’étendre à l’ensemble de la monocouche ou seulement par parcelles. Ils sont valorisés en kristas ou en points. Ainsi, si l'ensemble du monopole du CPU a subi un changement, il est estimé à 4 croisements ; si ¾ - de 3 cr ; ½ sur 2 cr; ¼ - pour 1 croix. Les formes de CPD dépendent des propriétés biologiques du virus, du type de cellules, de la dose d'infection, des conditions de culture, etc. Certains virus présentent une CPD après 2 à 3 jours, d’autres après 1 à 2 jours. 3 formes de CPU : fragmentation– destruction des cellules en fragments séparés, qui sont séparés du verre et passent dans le fluide culturel. Arrondi– les cellules perdent la capacité de s’attacher au verre, elles prennent une forme sphérique, se séparent et flottent librement là où elles meurent. Formation de symplastes– dissolution des membranes cellulaires, à la suite de laquelle les cytoplasmes des cellules voisines fusionnent, formant un tout unique dans lequel se trouvent les noyaux cellulaires. De telles formations sont appelées symplastes - cellules polyphages géantes. Il est nécessaire d'effectuer au moins 3 passages à l'aveugle afin de juger de la présence du virus dans le matériel testé. L'hémadsoption est la connexion des globules rouges avec la surface des cellules infectées par un virus.
51. Calcul du titre viral selon Reed et Mench. Titrage du virus avec effet évalué statistiquement avec calcul du titre par lecture et menu. Pour cette méthode de titrage, tout modèle biologique peut être utilisé, mais ce modèle doit être sensible au virus titré (cultures cellulaires, embryons, animaux de laboratoire). Selon l'effet infectieux des modèles biologiques infectés, ils sont répartis comme suit : cliniquement reconnus ; selon les changements pathomorphologiques ; au décès du modèle ; par l'accumulation d'hémagglutinine. Les résultats du travail dépendent de la dose du virus. Il a été établi que la dose du virus qui provoque 50 pour cent de l’effet infectieux est la moins sensible aux fluctuations et la plus déterminable de toutes les doses possibles. Le titre est exprimé en doses efficaces de 50 pour cent. Il s’agit d’une DE de 50. En fonction du modèle biologique utilisé et de l’effet obtenu, la dose de 50 pour cent peut être exprimée dans les unités suivantes : DL 50 – ID 50 DCE 50 AID 50 TsPD 50– il s’agit d’une dose cytopathogène de 50 pour cent déterminée dans des cultures cellulaires par CPD. Si dans les systèmes infectés nous n'observons pas 50 pour cent de l'effet ID 50, alors le titre est calculé par lecture et menu : lg LD 50 = lg ECD - (% années d'ECD - 50%) / (% années d'ECD - % années de DPE) TOUT CELA MULTIPLIÉ PAR lg multiplicité fois
36. Règles et horaires d'ouverture d'un laboratoire de virologie. Tous les étudiants sont instruits et formés à la sécurité. L'entrée dans les locaux de production par des personnes non autorisées, ainsi que l'entrée sans robe et sans chaussures de remplacement, est interdite. Il est interdit de sortir du laboratoire en blouse et casquette. Fumer, manger au laboratoire et conserver les aliments. Tout matériel entrant dans le laboratoire doit être considéré comme infecté. A la fin des travaux, le lieu de travail est remis en ordre et soigneusement anonymisé. Étiquetage des ustensiles contenant des matières infectieuses. Les mains portant des gants sont lavées dans un pot avec une solution de chloramide à 5 pour cent, puis les gants sont retirés et désinfectés une seconde fois, désinfectés et lavés. ra Le travail de virologue du laboratoire repose sur trois grands principes : prévenir l’infection des employés ou des personnes travaillant avec des matériaux contenant des virus. Eviter la contamination du matériel (les outils, les ustensiles sont stériles) en nettoyant les locaux avec une solution désinfectante + lampes ultraviolettes. Empêcher le virus de se propager à l'extérieur du laboratoire (avec l'air, les ustensiles, les matières solides et liquides). Les pipettes et les verres doivent être jetés dans le stérilisateur. Tubes à essai contenant des virus, des tissus - dans un autoclave. N'ouvrez pas la centrifugeuse jusqu'à ce qu'elle s'arrête. Il vous suffit d'éliminer l'air de la seringue à l'aide d'un coton-tige contenant 75 % d'alcool. Il est interdit d'aérer le local à l'aide d'un système de ventilation doté d'un filtre.
37. Précautions de sécurité avec le matériel contenant des virus. Empêcher la dispersion des virus dans l’environnement extérieur. Empêcher la contamination (contamination) du matériel contenant des virus par la microflore étrangère. Assurer la sécurité personnelle. Pour respecter ces exigences, les règles de travail suivantes sont nécessaires : être attentif et soigné ; être seulement en robe et se changer dans la garde-robe ; travailler uniquement avec des poignets boutonnés, une casquette et un masque de gaze ; maintenir strictement la propreté et l'ordre dans le laboratoire ; il ne doit y avoir aucun objet étranger sur le bureau ; Il est interdit de fumer et de manger. Utilisez des instruments et des ustensiles stériles. Travaillez avec des récipients proches de la flamme du brûleur. Ne mettez pas vos doigts dans votre bouche. Appareils usagés dans le stérilisateur. Recueillir les pipettes usagées dans un récipient contenant une solution désinfectante. Recueillir les déchets solides ou liquides (coton) dans des conteneurs spéciaux pour une désinfection ultérieure. Ne jetez pas de déchets dans les éviers ou les toilettes.
33. Le mécanisme de l'action antivirale de l'interféron. L'interféron n'a pas d'effet direct sur le virus. Elle affecte uniquement la cellule en activant la synthèse de certaines enzymes cellulaires. En particulier, l'enzyme protéine kinase et la 2,5 oligoasynthétase. Les informations sur la synthèse de ces enzymes se trouvent également dans certaines régions des gènes de la cellule et sont également dans un état répressif. Sous air, il y a dérépression des gènes responsables de la synthèse de la protéine kinase et de la 2,5 iligoAs synthétase. Et leur synthèse augmente fortement. 1) sous l'air de la protéine kinase, le facteur initiateur est phosphoré, ce qui assure la liaison de l'ARN messager viral au ribosome. Ainsi, l’ARN messager viral ne peut pas entrer en contact avec l’appareil ribosomal de la cellule, c’est-à-dire le début de la traduction. Et finalement, la synthèse des protéines et enzymes virales devient impossible. 2) sous l'influence de l'interféron, la synthèse de la 2,5 oligoAsynthétase est activée, ce qui catalyse la synthèse de l'acide 2,5 oligoadénylique dans la cellule. Cet acide modifie l’action des nucléases cellulaires pour détruire les ARN messagers viraux. Ainsi, sous l'influence de l'interféron, il se produit : un blocage de la traduction de l'ARN messager viral ; destruction des ARN messagers viraux. Effet inhibiteur de l'interféron sur la reproduction cellulaire : l'interféron à des concentrations de 0 à 1 000 unités par ml supprime la reproduction d'une grande variété de cellules dans n'importe quel tissu. L'interféron régule la croissance de nombreux types de cellules, notamment les cultures de cellules primaires et les cellules tumorales. Elle repose sur la suppression de la synthèse de certaines protéines cellulaires et la synthèse de nouvelles protéines par l'interféron. Inter augmente l'activité tueuse des lymphocytes T. À fortes doses, ils inhibent la formation d’anticorps. Au contraire, de petites doses stimulent la formation d’anticorps. Krilling – les cellules traitées avec de petites doses d’interf produisent plus d’interf que les cellules non traitées. Des doses trop élevées sont le processus inverse.
35. Types d'interactions entre le virus et la cellule. Productif et avorté. Productif est divisé en lytique et latent. Productif: Il s’agit d’un type d’interaction dans lequel une nouvelle génération de virion se forme dans la cellule. Si la cellule meurt rapidement après avoir acquis un nouveau virion, il s'agit alors d'une voie lytique productive d'interaction entre le virus et la cellule. Si la cellule dans laquelle le virus se développe pendant longtemps conserve sa viabilité (par bourgeonnement), il s'agit alors du produit d'une interaction de type latente. Abortif: Ce type est réciproque lorsque la reproduction des virions s'arrête à n'importe quel stade, le virion ne se développe pas. À la suite de l'interaction du virus avec la cellule, les changements suivants peuvent se produire dans la cellule : dégénérescence cellulaire– les cellules se transforment d'abord en une forme irrégulière, puis s'arrondissent, une granularité apparaît dans le cytoplasme, puis une fragmentation nucléaire puis la mort cellulaire. De tels changements sont appelés CPD. En croisements : 4 croisements – 100% d’efficacité. Formation de symplastes– des cellules multinucléées. Formation d’organismes d’inclusion– contenant du mb intranucléaire et plasmatique, de l'ARN ou de l'ADN. Transformation cellulaire– les virus oncogènes (rétrovirus à ARN). La reproduction de virus oncogènes dans une cellule ne s'accompagne pas de CPD. La cellule produit constamment un virus. Synthèse d'interféron.
4. Résistance des virus aux facteurs physico-chimiques. La résistance des virus animaux a été relativement bien étudiée lorsqu'ils sont exposés à des facteurs externes : température, rayonnements, ultraviolets, ultrasons, pH, formaldéhyde, phénol, etc. Pour se protéger de ces influences, les virions possèdent une coque protéique. La structure et la composition chimique différentes des enveloppes protéiques déterminent la stabilité variable des virus. En fonction de ces caractéristiques, le même facteur peut détruire complètement certains virions et pas d’autres. Par exemple, les solvants organiques : les virions dans les coquilles desquels il n'y a pas de lipides sont résistants à ces substances, et ceux contenant des lipides sont rapidement détruits. L'inactivation des virus signifie la perte totale ou partielle de leur activité biologique, résultant de l'action de facteurs physiques et chimiques. Lorsque l'acide nucléique et la protéine virale changent, une inactivation complète se produit, c'est-à-dire une perte de toutes les propriétés biologiques du virus - il perd uniquement ses propriétés infectieuses et conserve son immunogénicité. La nature et l'étendue des agents de nature chimique et physique agissant sur le virus dépendent de la nature du facteur inactivant, de la dose, pendant longtemps, du type de virus. Lorsque le virus est inactivé, soit le clivage des protéines de l'enveloppe peut se produire, suivi de sa désintégration en unités séparées, soit le compactage des protéines tout en maintenant la structure globale de l'enveloppe. Le clivage est observé sous l'action d'un environnement acide et alcalin avec un chauffage prolongé et faible. La coagulation et le compactage se produisent lorsqu'ils sont exposés au formaldéhyde, à une température élevée ou au phénol. Cela dépend de la concentration et de la durée. Ainsi, dans certains cas, la coagulation des protéines s’accompagne de la destruction des acides nucléaires et le virus subit une perte irréversible de son pouvoir infectieux. Dans d’autres cas, la capacité du virus à se reproduire est préservée. Conservé avec de la glycérine.
60. RAP. Le principe de la méthode : un gène spécifique à un virus donné est identifié - une section d'une molécule d'ADN qui transporte des informations pour la synthèse d'une protéine. Ce gène est ensuite identifié dans le matériel de test par PCR. Cette réaction permet la formation de copies supplémentaires du gène - amplification d'une section d'ADN dans un tube à essai. Selon le but de l'étude, l'espèce ou le genre de mo peut être identifié. L'essence de la PCR : la molécule d'ADN est chauffée à 90-94 degrés. Ce qui conduit à la destruction des liaisons hydrogène entre les bases azotées de la double hélice puis refroidie à 52 g en présence de l'enzyme ADN polymyrase. Une augmentation ultérieure du taux conduit à la synthèse d'une nouvelle molécule d'ADN - une matrice complémentaire. Cette procédure est répétée plusieurs fois, ce qui donne des fragments plus gros. L'indication est réalisée par électrophorèse ou par sonde ADN marquée. Principaux composants : l'ADN polymyrase est thermostable ; oligonucléotide de 20 nucléotides ; les triphosphates ; amplificateur, verrerie et réactifs pour électrophorèse en gel d'agarose. Mise en place : obtention d'un échantillon d'ADN. Pour ce faire, le matériau étudié est mis en suspension dans un tampon ou de l'eau distillée. Ajoutez du sodium OH et maintenez pendant 7 minutes. Le mélange est neutralisé. Le lysat est centrifugé pendant 10 minutes pour sédimenter les grosses particules. Le liquide surnageant est utilisé pour la PCR. La PCR est l'amplification d'un gène donné d'un fragment d'ADN. Ensuite, il est fondu dans un thermocycleur pendant 3 heures. Indication d'amplification - l'échantillon est soumis à une électrophorèse dans un gel d'agarose pour séparer l'ADN. Au bout de 30 minutes, l'agarose est polymisé dans l'appareil et des trous se forment dans l'agarose. 10 µl du mélange sont prélevés et mélangés à 5 µl de colorant. Le mélange est ajouté dans les puits et une électrophorèse est réalisée pendant 40 minutes. La plaque est retirée et colorée dans une solution de bromure pendant 10 minutes. L'agarose est ensuite placé sur un transilluminateur et les motifs de bandes résultants sont photographiés. Les bandes révélées par le rayonnement ultraviolet sont des fragments d'ADN.
49. Méthode d'infection des cultures cellulaires. Indication des virus dans les cultures cellulaires. Infection : à cet effet, des tubes avec une monocouche cellulaire continue sont sélectionnés. Le milieu de croissance est drainé et les cellules sont lavées plusieurs fois avec la solution de Hank. 0,2 à 0,1 ml de matériel viral est ajouté à chaque tube et réparti uniformément sur toute la couche de cellules par agitation. Sous cette forme, les tubes sont laissés 1 à 2 heures à 22 ou 37 degrés pour l'adsorption du virus à la surface des cellules. Ensuite, le matériel viral est retiré des tubes à essai et le milieu d'entretien est versé dans le tube à essai (1-2 ml). Après isolement du virus, la monocouche de cellules est lavée 2 fois avec la solution de Hank puis le milieu de support est versé. Indication: selon DPC ; RGAd ; par formation de plaque ; inclusions intracellulaires; RÉCIF; microscopie électronique
54. RTGA. RGA. RTGA : essence– lorsque le virus est mélangé à un sérum spécial, le virus perd ses propriétés hémagglutinantes. Objectifs– identification du vir isolé ; détection des anticorps dans le sérum testé et de leur titre. Composants– pour le sérovariant direct : matériel viral, sérum spécifique, suspension d'érythrocytes à 1 %, solution saline pour dilution. Pour rétrospective - sérum de test, antigène standard dans une certaine dose de 4 GAE (titre de dilution du virus) 4 GAE - 1:32. Schème– à chaque dilution de sérum ajouter un volume égal d'antigène étalon (virus) à la dose de 4 GAE. Contacter 30 minutes à température ambiante. Dans chaque puits contenant des dilutions de sérum et une dose constante de virus sur 4 ha, ajouter un volume égal de suspension de globules rouges. Contacter 30 à 60 min à température ambiante. Comptabilité les réactions s'effectuent en crêtes. si c’est un plus, alors il n’y a pas d’agglutination ; si c’est quelques minutes, alors c’est une hémagglutination. Le titre d'anticorps dans le sérum testé est la dilution maximale du sérum qui retarde complètement l'agglutination des globules rouges.RGA : essence : dans l’adsorption du virus à la surface du globule rouge, ce qui conduit au collage. Finalités : indication ; pour le titrage des virus à Haen. Composants : virus ; 0,5 suspension de globules rouges ; solution saline pour la préparation. Schéma post : préparer une dilution en deux du virus ; ajouter un volume égal de suspension de globules rouges à 0,5 % à chaque développement de virus ; contacter 30 à 60 minutes à température ambiante. Comptabilité : en cristaux. 4 crêtes – 100 % d’agglutination. 3 cristaux - 75% . 1 croix – agglutination. 1 haen est la dilution maximale du virus pouvant provoquer une agglutination de 50 % des globules rouges.
57. ELISA. L'essence : lorsque l'antigène + le sérum marqué se lie, l'enzyme décompose le substrat. Un complexe antigène + conjugué est formé pour former un produit de réaction coloré, évalué au microscope optique ou visuellement. Objectif : identification. Composants : matériau de soude virale, conjugué, substrat. Schéma de prise : la cellule culitra est fixée avec de l'acétone refroidie. Ils sont séchés et le conjugué leur est appliqué. Incuber pendant 1 à 2 heures à une température de 37 degrés dans une chambre humide. Laver avec une solution saline, rincer à l'eau distillée et sécher. Quelques gouttes de la solution de substrat y sont appliquées, incubées pendant 5 à 10 minutes, puis lavées dans une solution saline et rincées à l'eau claire. Comptabilité : dans ce cas, c'est-à-dire en présence d'antigène, après application du conjugué, il se forme un complexe antigène plus anticorps marqué avec une enzyme. Après application du substrat, celui-ci se décompose sous l'action d'une enzyme, formant un produit coloré bien visible au microscope optique.
56. RSK. L’essence : la liaison d’un complément au complexe antigène plus anticorps. L'absence de complément libre dans ce système est jugée par la rétention d'hémolysine dans le système indicateur. Objectifs : identification ; détection des anticorps et de leur titre dans le sérum sanguin à tester. Composants : 2 systèmes – 1 (matériel contenant le virus ; sérum spécifique ;) (sérum de test ; antigène standard). 2) système hémolytique (indicateur) - une suspension à 2-3 % d'érythrocytes de mouton est un antigène ; l'hémolysine (sérum hémolytique) est un anticorps. Les anticorps correspondent à l'antigène. Et un compliment pour 1 seule réaction : si c'est le premier, il y aura un retard dans l'hémolyse lorsque le complément entrera en contact avec le système étudié. Si au deuxième, les globules rouges sont lysés, il y aura une hémolyse complète. Schéma de configuration : la réaction est d'abord réalisée dans le système étudié, puis le système indicateur est ajouté dans le même tube à essai. Comptabilité : RSC positif – hémolyse retardée. Négatif – hémolyse complète.
7. Protéines virales. Constitué d'acides aminés. La composition de la protéine virale dépend de l'ordre d'alternance des acides aminés ; cet ordre est déterminé par l'information génétique contenue dans le génome viral. Les protéines virales sont divisées en protéines structurelles et non structurelles. Les protéines structurelles font partie des virions matures. Les b non structurels ne sont pas inclus dans les virions matures mais sont obligatoires à certains stades de la reproduction. De constructionNon structurel
8. Enzymes virales. Ils sont de nature protéique. Ils peuvent être associés directement au virion, mais ils ne sont pas associés – ni structurels. dans l'ADN Pour l'ARN : ADN polymyrase ARN-dépendant - il n'est pas présent dans la cellule, il est nécessaire pour "-" contenant de l'ARN et de l'ADN, qui dans la famille vir des rétroviridae contient une enzyme qui transduit le génome viral est appelée ADN polymyrase ARN-dépendant. Cette enzyme porte les noms : révertase, transcriptase inverse. Enzymes impliquées dans la formation des protéines virales : protéases, protéine kenase.
52. Infirmière autorisée.Essence: Lorsque le virus interagit avec un sérum spécifique, le virus perd ses propriétés infectieuses, sa capacité à se multiplier dans les cellules. Objectifs: identification du virus isolé, détection des anticorps dans le sérum sanguin et titre d'anticorps. Composants: matériel contenant des virus, sérum spécifique, modèle biologique. Si rétrospectif : test sérum sanguin, antigène standard, modèle biologique. Schéma de configuration générale: mélanger l'antigène et l'anticorps, contact pendant 30-40 minutes, maximum 2 heures à une température de 37-38 degrés ; un mélange d'antigène et d'anticorps est utilisé pour infecter un modèle biologique ; observation et comptabilité. Comptabilité: Un pH positif signifie vivant, un pH négatif signifie mort.
53. RDP.Essence: les mêmes antigène et anticorps placés à même distance l'un de l'autre dans un gel d'agar diffusent l'un vers l'autre, formant un précipité sous forme d'une bande blanche au point de rencontre. Objectifs: identification du virus isolé, détection des anticorps dans le sérum à tester. Composants: matériel contenant des virus, sérum spécifique, gel d'agar. Pour une rétrospective : testez le sérum sanguin, l'antigène standard, le gel d'agar. Schéma de mise en scène : des revêtements de gélose sont préparés sur une lame de verre, des puits sont préparés, des composants de réaction sont ajoutés aux puits selon un certain schéma, le verre avec la réaction est placé dans un thermostat à 37-38 C. La réaction est enregistrée après 48 heures. Une réaction positive est la formation d’une bande de précipitation blanche.
55. RGAd, RTGAd. RGAd : Essence: dans l'adsorption des érythrocytes à la surface des cellules infectées par le virus. Objectifs: indication virale. Composants: culture cellulaire infectée par du matériel contenant un virus ; suspension de globules rouges . Schéma de mise en scène : pré-infecter une culture cellulaire monocouche avec le matériel de test. Les cultures sont drainées dans un milieu de culture de support. Laver avec la solution Hanks. Une suspension de globules rouges est ajoutée. Contacter 5 à 15 minutes à température ambiante. Comptabilité: réalisée au microscope optique. Positif – les globules rouges sont adsorbés sur les cellules ; négatif – les globules rouges flottent librement. RTGAd : essentiel: dans la liaison d'anticorps spécifiques à la surface des cellules infectées par le virus, ce qui conduit à l'inhibition de l'adsorption sur les cellules érythrocytaires. Objectifs: identification du virus isolé. Composants: culture cellulaire contaminée; sérum spécifique ; suspension de globules rouges. Schéma de mise en scène : une culture cellulaire monocouche est pré-infectée avec un matériau initial monocouche contenant un virus. Verser dans un milieu nourrissant et ajouter 0,8 ml de sérum spécifique. Contactez 20-30 minutes. Une suspension de globules rouges est ajoutée. Contacter 5 à 15 minutes. Comptabilité: Pour le contrôle, ils doivent installer RGA. Comptabilisation dans les tubes expérimentaux : positif - les globules rouges flottent librement, négatif - les globules rouges flottent également librement. Comptabilisation dans des tubes de contrôle avec positif – adsorption, négatif – flottement libre.
6. Acides nucléiques viraux.+ L'ARN est un acide nucléique viral qui a également la fonction d'ARN informatif. Les informations sur le système de synthèse des protéines dans l’ARN+ sont immédiatement transférées à l’ARN génomique sans transcription. -Les virus contenant de l'ARN sont des virus avec un ARN simple brin qui n'a pas la fonction d'ARN messager ; dans ces virus, la synthèse de l'ARN messager (transcription) se produit sur la matrice moins les brins d'ARN génomique en utilisant une enzyme spécifique du virus étroitement associée. avec ARN gnome, ARN polymyrase ARN-dépendante. Il existe des virus contenant à la fois des brins d’ARN plus et moins, notamment les adénovirus et les paramyxovirus. Les informations génomiques dans l'ADN double brin sont codées sur les deux brins. Les acides nucléiques sont représentés par des polynucléotides constitués de nucléotides individuels. Leur quantité en acide nucléique varie. Chaque nucléotide est constitué de 3 sous-unités : un résidu d'acide phosphorique, un glucide et une base azotée.
9. Structure des virus. Formulaires de base. Types de symétrie. Structure : ADN : généralement double brin, les informations génétiques sont codées sur les deux brins. L’ADN viral peut être disposé de manière linéaire et circulaire. Peut être monocaténaire. ARN viraux : souvent simple brin, moins souvent double brin. Disposé linéairement, de manière circulaire, fragmenté. En règle générale, ils se composent de 11 à 12 fragments. L'ARN vir simple brin peut être de deux types : les brins plus et les ARN brins moins (génome négatif) Types de symétrie : l'emplacement des sous-unités protéiques (caposmères) détermine le type de symétrie du virion - hélicoïdale, cubique, combinée. Spirale Il s’agit d’un type de simulation dans lequel les capsomères sont situés autour de l’acide nucléique de manière hélicoïdale. Les grands virus et certains virus de taille moyenne ont ce type de simulation. Forme : en forme de bâtonnet, en forme de poly, sphérique, ovale. Dans les virus en forme de bâtonnet, la capside est constituée de capsomères disposés autour de l'acide nucléique en spirales de même diamètre, étroitement adjacentes les unes aux autres. Dans les virus sphériques, les capsomères sont disposés en spirale mais de diamètres différents. Type cubique : La plupart des petits virus et une proportion importante de virus de taille moyenne en sont atteints. La forme de ces virus est sphérique. Les capsomères de la capside sont situés autour des noyaux acides comme autour d'un corps isométrique régulier. L’enveloppe protéique de ces virus se rapproche de la forme d’un icosaider, un visage régulier à 20 côtés. Combiné type de symétrie : se compose de spirale et de cube. Tous les phages et certains virus complexes de la famille des coxviridae en sont dotés. Ils ont une coque externe cubique et une coque capside en spirale. Les phages ont une tête icoseindrique et un processus en spirale.
18. Les principales étapes de la première phase de reproduction virale. C'est la phase d'infection de la cellule, durant cette phase le virion doit entrer en contact avec la cellule, pénétrer dans la cellule et se déshabiller. D'abord l'étape d'adsorption des virions à la surface cellulaire peut se dérouler de deux manières : physico-chimique (non spécifique) ; récepteur (spécifique). La voie physicochimique est déterminée par l’interaction des forces électrostatiques de surface qui apparaissent entre les groupes chargés positivement de protéines virales et les groupes carboxines, sulfates et phosphates chargés négativement de la paroi cellulaire. Récepteur basé sur l'interaction spécifique du récepteur de la protéine virale avec des récepteurs complémentaires à la surface de la paroi cellulaire. Les récepteurs des virus et les récepteurs des cellules sensibles à un virus donné ont une configuration complémentaire (comme la clé d'une serrure). Si la cellule n’est pas sensible, la réabsorption ne se produira jamais. Deuxième pénétration – se produit de différentes manières pour différents virus : à l’aide de viropexys ; en fusionnant les coquilles. Viropexie– cette voie est similaire à la pinocytose. Tout d'abord, au site d'adsorption à la surface cellulaire, une invagination de la paroi cellulaire de la membrane se produit, puis les bords de la membrane se ferment avec l'intérieur de la cellule, le virion avec toutes ses membranes apparaît dans la vacuole cellulaire. Par la fusion- dans ce cas, les zones de l'enveloppe virale et de la membrane cellulaire qui se reçoivent fondent sous l'action d'enzymes spécifiques du virus et seul l'acide nucléique viral apparaît dans la cellule, tandis que les restes du virus s'accumulent dans la cellule. membrane cellulaire. Troisième scène– la déproténation – libération des membranes – dépend des voies d’entrée du virus dans la cellule. Si la déprotonisation n’est pas isolée en une étape distincte par fusion des membranes, elle se produit simultanément à la pénétration du virus. Si la pénétration se fait par viropexie, alors la libération de l'acide nucléique viral des enveloppes commence après la destruction des protéines, des lipides et des graisses qui composent les enveloppes virales. Toutes les étapes dépendent de la température.
20. Transcription. Il s’agit de la réécriture de l’information génétique de l’acide nucléique viral en ARN d’information virale, nouvellement synthétisé selon les lois du code génétique. (le virus doit présenter la protéine à la cellule en cours de synthèse et être convertie en ARN). Le produit final de la transcription est l’ARN messager viral. Les ARN + simple brin n'ont pas de transcrits, mais leur ARN viral génomique contient l'information vir ARN. Dans l'ARN-simple brin, le génome ne peut pas remplir la fonction d'ARN messager et son ARN est transflexé à l'aide de l'enzyme ARN polymyrase ARN-dépendante spécifique du virus. DESSIN!
21. Diffusion. Il s’agit du processus de traduction de l’information génétique contenue dans les ARN messagers viraux en une séquence d’acides aminés spécifique. Une traduction se produit lorsque quatre bases intégrées dans l’ARN messager viral sont converties en un code de 20 acides aminés. Le produit final de la traduction est constitué de protéines virales. La synthèse des protéines se produit sur les ribosomes cellulaires. Se compose de 3 phases : initiation de la traduction et début de la traduction ; continuation; terminaison – fin de la diffusion. L'initiation repose sur la formation d'un complexe de composants nécessaires au début de la traduction, c'est-à-dire que le complexe d'initiation repose également sur la reconnaissance du ribosome par l'ARN messager viral et sa liaison à certaines zones appelées coiffe. C'est de la guanine méthylée. Après avoir reconnu le capuchon, le ribosome glisse le long de la molécule d'ARN messager jusqu'à ce qu'il atteigne le site où commence le décodage de l'information.
5. Composition chimique des virus. Les virus sont constitués de acides nucléiques (ADN, ARN). Les acides nucléiques sont représentés par des polynucléotides constitués de non-nucléotides individuels. Chaque nucléotide est constitué de 3 sous-unités : un résidu d'acide phosphorique, un glucide et une base azotée. Protéines virales : Composé d'acides aminés. La composition de la protéine virale dépend de l'ordre d'alternance des acides aminés ; cet ordre est déterminé par l'information génétique contenue dans le génome viral. Les protéines virales sont divisées en protéines structurelles et non structurelles. Les protéines structurelles font partie des virions matures. Les b non structurels ne sont pas inclus dans les virions matures mais sont obligatoires à certains stades de la reproduction. De construction En fonction de leur emplacement dans le virion, les protéines virales sont divisées dans le groupe suivant : protéines de capside - dans la capside ; supercapside b – dans la supercapside (principalement des protéines, il y a aussi des graisses et des glucides) ; protéines matricielles – protéines de la couche membranaire ; les protéines du noyau viral sont représentées par des enzymes. Non structurel– selon les fonctions qu'ils remplissent, ils se répartissent en : régulateur de l'expression du génome viral ; des inhibiteurs de biosynthèse cellulaire ; des inducteurs de destruction cellulaire ; précurseurs de protéines virales, protéines vir structurelles ; certaines enzymes virales ne font pas partie des virions matures. Lipides: sont principalement inclus dans une partie de la coquille surercapside du virion dans les virus complexes. Tous ne sont pas codés par le génome vir et sont d’origine cellulaire. Ils sont représentés par les phospholipides et les glycolipides. Les glucides: font partie de la supercapside obole, ne sont pas codées par le génome viral et sont d'origine cellulaire, représentées par des glycoprotéines et des glycolipides . Enzymes virales : Ils sont de nature protéique. Ils peuvent être associés directement au virion, mais ils ne sont pas associés – ni structurels. Les enzymes polymyrases précoces et réplicases précoces participent à l’étape de changement d’information. Ils sont classés comme inhibiteurs de la biosynthèse cellulaire. Enzymes transhibant le génome viral : dans l'ADN contenant des virus - Des ARN polymyrases ADN-dépendantes sont présentes dans la cellule ; dans certains cas, elles sont accessibles aux virus, dans d'autres, elles ne le sont pas. Elle peut être d’origine cellulaire ou virale. Les virus contenant de l'ADN qui se reproduisent dans le noyau sont d'origine cellulaire. Dans le cytoplasme - origine virale - spécifique au virus.
Pour l'ARN : ADN polymyrase ARN-dépendant - il n'est pas présent dans la cellule, il est nécessaire pour "-" contenant de l'ARN et de l'ADN, qui dans la famille vir des rétroviridae contient une enzyme qui transduit le génome viral est appelée ADN polymyrase ARN-dépendant. Cette enzyme porte les noms : révertase, transcriptase inverse. Enzymes impliquées dans la formation des protéines virales : protéases, protéine kenase.
13. Bactériophages. Virus bactérien. Les bactériophages à ADN et à ARN sont connus. La plupart de l’ADN des phages est double brin. Les phages à ARN sont simple brin. L'acide nucléique du phage est entouré d'une capside polyédrique (tête), à laquelle, dans de nombreux phages, un appendice (queue) est attaché. Le diamètre des têtes est d'environ 60 à 95 nm et la longueur des processus est de 250 nm avec une épaisseur de 10 à 25 nm. Le processus sert de structure d’attachement à la bactérie. L'interaction entre les cellules b et microbiennes est un processus biologique complexe dont l'issue dépend des propriétés des phages et se manifeste par la lyse des cellules bactériennes. les bactériophages sont utilisés pour le diagnostic (anthrax) ; pour le traitement des infections bactériennes ; pour la prévention de l'inf (salmonellose). DESSIN!
22. Réplication de l'ADN viral.
23. Réplication de l'ARN viral. ARN simple brin avec un génome négatif :
25. Assemblage des virions et leur libération de la cellule.
: explosion, rupture, destruction de la cellule dans laquelle se sont formés des virions matures (virus simples), la cellule meurt. Les structures complexes naissent par bourgeonnement, c'est-à-dire qu'elles sortent de la paroi cellulaire et bourgeonnent. Dans ce cas, la cellule ne meurt pas immédiatement, mais lorsque ses réserves sont épuisées
19. Deuxième phase de reproduction.Étape éclipse :étape de changement d’information. À ce stade, la fonction du génome cellulaire est supprimée du fait que l’acide nucléique bloque le virus et que les enzymes virales bloquent l’appareil génétique de la cellule et les systèmes de synthèse de la cellule. Cela amène la cellule à cesser de reproduire ses propres composants cellulaires et à passer à la reproduction de composants externes. Les premières réplicases et les premières polymyrases sont impliquées ici. Transcription: Il s’agit de la réécriture de l’information génétique de l’acide nucléique viral en ARN d’information virale, nouvellement synthétisé selon les lois du code génétique. (le virus doit présenter la protéine à la cellule en cours de synthèse et être convertie en ARN). Le produit final de la transcription est l’ARN messager viral. Les ARN + simple brin n'ont pas de transcrits, mais leur ARN viral génomique contient l'information vir ARN. Dans l'ARN-simple brin, le génome ne peut pas remplir la fonction d'ARN messager et son ARN est transflexé à l'aide de l'enzyme ARN polymyrase ARN-dépendante spécifique du virus. DESSIN!Diffuser: Il s’agit du processus de traduction de l’information génétique contenue dans les ARN messagers viraux en une séquence d’acides aminés spécifique. Une traduction se produit lorsque quatre bases intégrées dans l’ARN messager viral sont converties en un code de 20 acides aminés. Le produit final de la traduction est constitué de protéines virales. La synthèse des protéines se produit sur les ribosomes cellulaires. Se compose de 3 phases : initiation de la traduction et début de la traduction ; continuation; terminaison – fin de la diffusion. L'initiation repose sur la formation d'un complexe de composants nécessaires au début de la traduction, c'est-à-dire que le complexe d'initiation repose également sur la reconnaissance du ribosome par l'ARN messager viral et sa liaison à certaines zones appelées coiffe. C'est de la guanine méthylée. Après avoir reconnu le capuchon, le ribosome glisse le long de la molécule d'ARN messager jusqu'à ce qu'il atteigne le site où commence le décodage de l'information. Réplication de l'ADN viral : Réplication de l'ADN double brin : Une molécule d'ADN double brin est d'abord séparée en 2 brins distincts à l'aide d'enzymes nucléases cellulaires, puis l'ADN d'information virale est formé sur l'un des brins d'ADN viral dont est la matrice. Cela se produit à l’aide d’une enzyme ARN polymyrase ADN-dépendante spécifique du virus ou cellulaire. Ensuite, l’ARN informatif viral se déplace vers le ribosome cellulaire où la traduction se produit avec la formation de protéines et d’enzymes virales, y compris l’enzyme ADN polymyrase. Grâce à l'ADN polymyrase, un deuxième brin complémentaire d'ADN est construit à partir des nucléotides cellulaires. De cette manière, de nouvelles molécules d’ADN double brin sont synthétisées. Réplication de l'ADN simple brin : Les simples brins d’ADN ont une polarité positive. À l’aide de l’ADN polymyrase ADN-dépendante, une enzyme spécifique du virus, un brin négatif complémentaire d’ADN est formé sur la matrice d’ADN viral simple brin. Une structure en double hélice est synthétisée, appelée forme réplicative. Ensuite, sur la matrice, les brins négatifs de la forme réplicative forment des brins plus d’ADN simple brin en déplaçant les brins plus d’ADN de la forme réplicative. Réplication de l'ARN viral : ARN simple brin avec un génome négatif : Immédiatement après la pénétration dans la cellule, la transcription se produit avec la formation d'ARN inf viral plus. L’ARN polymyrase ARN-dépendante, enzyme spécifique du virus, est impliquée dans ce processus. L'ARN Vir inf est ensuite traduit pour former des protéines et l'enzyme ARN polymyrase. Par la suite, à l'aide de l'ARN polymyrase, des brins d'ARN simples brins filles moins sont formés sur la matrice ainsi que des brins d'ARN d'information. ARN simple brin avec un génome positif : Après pénétration dans la cellule, l'ARN plus se lie immédiatement aux ribosomes où il est traduit pour former des protéines et des enzymes, notamment l'enzyme ARN réplicase. Ensuite, sous l'action de l'ARN réplicase, une forme réplicative se forme. Sur la matrice, l'ARN moins de la forme réplicative crée une image de brins d'ARN plus en les déplaçant de la forme réplicative. Réplication de l'ARN viral double brin : la synthèse des ARN viraux messagers se produit sur une matrice d'ARN double brin en utilisant l'enzyme ARN polymyrase ARN-dépendante. Transcription sur une matrice de brin d'ARN, chaque fragment est transcrit séparément. Ensuite, ils sont traduits pour former des protéines et l'enzyme ARN polymyrase à l'aide de cette enzyme sur les brins positifs de l'ARN messager pour former des brins négatifs d'ARN complémentaires, c'est à dire de l'ARN double brin. Assemblage des virions et leur libération de la cellule : 2 stratégies d'assemblage, de maturation et de sortie d'une cellule infectée : mise en œuvre de l'assemblage et de la maturation au sein des cellules ; combinaison de la dernière étape de l’assemblage du virion avec la sortie de la cellule infectée.
L'assemblage s'effectue par simple agrégation, c'est-à-dire que la combinaison d'une protéine vir avec un acide nucléique se produit sous l'influence de facteurs physico-chimiques, c'est-à-dire qu'un auto-assemblage se produit. Il est basé sur l'unification et la reconnaissance spécifique des acides non protéiques et protéiques-nucléiques. Une nucléocapside se forme. Pour les virus simples, c’est là que se termine le processus d’auto-assemblage. Dans les virus complexes, le processus d’auto-assemblage s’effectue différemment. Une partie de la protéine va à la formation de la nucléocapside, qui se forme comme dans les virus simples, et une partie des protéines se déplace vers la membrane cellulaire. La nucléocapside formée s'y déplace ensuite. Et la formation d'une coque supercapside se produit lorsque le virion quitte la cellule, c'est-à-dire que la nucléocapside est recouverte de protéines qui se sont déplacées vers la membrane cellulaire et, à la sortie, les graisses et les glucides de la cellule sont intégrés dans cette coque externe. . Il y a deux façons de sortir: explosion, rupture, destruction de la cellule dans laquelle se sont formés des virions matures (virus simples), la cellule meurt. Les structures complexes naissent par bourgeonnement, c'est-à-dire qu'elles sortent de la paroi cellulaire et bourgeonnent. Dans ce cas, la cellule ne meurt pas immédiatement, mais lorsque ses réserves sont épuisées.
62-63. Variale ovine et caprine(genre Caprippoxvirus). Ecthyma contagieux des moutons et des chèvres(genre Parapoxvirus). Famille : poxviridae. Contenant de l'ADN. Caractéristiques de la reproduction : le génome du virus est très volumineux. Même dans les cellules infectées, la réplication est terminée au bout de 6 heures. La pénétration se produit par fusion des membranes virales et cellulaires. Après pénétration, l’ADN double brin est divisé et la réplication commence sur les deux brins d’ADN en même temps. De plus, la synthèse des composants viraux se produit dans le cytoplasme des cellules. Assemblage du virion dans le cytoplasme. Sortez en bourgeonnant.
2. Le rôle des virus dans la pathologie infectieuse des animaux. Actuellement, les maladies virales revêtent une grande importance dans la pathologie infectieuse des animaux, des humains et des plantes. Leur rôle augmente avec la réduction et l'élimination des maladies bactériennes, mycosiques et protozoaires. Les maladies virales représentent environ 80 pour cent en médecine et 50 pour cent en médecine vétérinaire. Il existe plus de 500 maladies connues causées par des virus. L'origine virale a été établie dans des maladies particulièrement dangereuses comme la fièvre aphteuse, la peste bovine, la peste porcine, etc. La virologie peut être divisée en générale et spécifique. Le général étudie la nature et l'origine des virus, leur classification, leur structure et leur composition chimique, la génétique et la sélection, les méthodes de diagnostic et de prévention, les bases de l'immunité antivirale. Le vir privé étudie le nom et la position systématique d'agents pathogènes spécifiques, la structure, la taille et la stabilité du virion, la thérapie, les méthodes de diagnostic et la prévention.
1. Histoire du développement. La première période commence depuis l’Antiquité jusqu’en 1892. Durant cette période, la virologie en tant que science indépendante n’existait pas. Les bactériologistes ont étudié des maladies d'étiologie inconnue. La deuxième période – la formation de la virologie en tant que science elle-même – couvre la période 1892-1950. cette période a commencé avec la découverte du botaniste russe D.I. Ivanovsky (1898) sur la filiabilité de l'agent causal de la mosaïque du tabac. Ivanovsky, étudiant l'étiologie de la maladie du tabac, a établi que cette maladie est causée par un petit micro-organisme spécial qui traverse des filtres bactériens. Il est invisible au microscope optique. Ne pousse pas sur des supports de croissance artificiels. Par la suite, des MO similaires ont été isolés d’autres plantes, ainsi que d’animaux et d’humains. Ils ont été réunis en un groupe indépendant : les ultravirus. Dans les années 1930, les embryons de poulet ont commencé à être utilisés en pratique virologique. En 1956, Stanley réussit à diviser le virus en ses principaux composants : les protéines et l'acide nucléique. À la fin des années 40 du 20e siècle, création des micros électroniques Rudenberg. Et la lumière a été créée par Leeuwenhoek. Scientifiques soviétiques ayant contribué à la virologie : Zhdanov, Likhachev, Syurin.
48. Méthode d'obtention de cultures cellulaires trypsinisées primaires monocouches. Des cellules monocouches sont nécessaires pour indiquer la présence du virus. Il est obtenu à partir de tissus ou d'organes en les traitant avec de la trypsine. Les monocouches sont divisées en 4 groupes : primaire trypsinisé ; repiquages ; diploïde ou semi-greffé ; greffé. Le tissu est écrasé et dispersé avec l'enzyme trypsine. Ensuite, la trypsine est éliminée par centrifugation et un certain volume de milieu nutritif liquide est ajouté au sédiment obtenu. Les cellules sont cultivées en une seule couche, une monocouche, sur la surface interne du verre. Il existe un besoin constant d’organes provenant d’animaux sains. ET des embryons âgés de 9 à 112 jours sont utilisés. Ovoscopie. Traitement des coquilles. À l'aide de ciseaux, coupez la coquille au-dessus du bord du puga. L'embryon est retiré stérilement. Laver avec la solution de Hank. Un sac cutanéo-musculaire est préparé. Laver avec la solution de Hank. Le tissu est déchiqueté avec des ciseaux. Transférer dans un flacon de trypsinisation. Le flacon est placé sur un agitateur magnétique pendant 15 minutes. La suspension est refroidie dans un récipient avec de la glace. Filtrer dans un flacon récepteur. La suspension cellulaire est centrifugée pendant 10 à 15 minutes. La trypsine est supprimée. Une masse combinée est préparée à partir du sédiment cellulaire, versée dans des tubes de 1 ml et les cellules sont cultivées.
47. Solutions de base et milieux nutritifs.Par origine Il existe des supports d’alimentation naturels et des supports d’alimentation artificiels. Produits naturels - issus de produits biologiquement actifs : vitalité embryonnaire et allantoïdienne ainsi que ajout de solutions salines équilibrées. Artificiel - préparé à partir de composants individuels. Le plus souvent, des supports d'alimentation universels sont utilisés, ou il peut y en avoir des spéciaux. Universal est le médium 199 et le médium Igla. La composition des milieux artistiques doit comprendre des acides aminés, des vitamines, des enzymes et des solutions salines équilibrées, parfois un indicateur (rouge de phénol). L'essence de l'indicateur est de détecter le virus en changeant de couleur. Au cours de la vie de la cellule, le pH évolue vers le côté acide. Dans un environnement acide, la couleur de l'indicateur passe du rouge cramoisi au jaune. Du sérum sanguin normal est parfois ajouté au milieu nutritionnel dans un volume représentant 100 pour cent du volume du milieu nutritionnel. Le sérum sanguin est appelé facteur de croissance. Il est ajouté pour la prolifération cellulaire, uniquement dans les milieux de croissance . Par finalité d'utilisation : milieux de croissance - sérum inclus ; soutenir – pas de sérum. Solutions salines équilibrées : toutes sont des dérivés de solution saline. Utilisé comme base pour la préparation des milieux de fosse et pour toutes manipulations de cultures cellulaires (pour laver quelque chose). Ce sont les solutions Hanks et Earle. Solutions dispersantes : pour séparer les cellules des autres et les cellules du verre. Solutions de pepsine, trypsine. Du verre - solutions versines. La solution de versine lie les cations calcium.
50. Titre de virus. Le titre de virus est la quantité de virus, c’est-à-dire la dose par unité de volume de matière contenant le virus. 3 méthodes de titrage : 1 méthode : titrage du virus en fonction de l'effet infectieux du virus avec un effet statistiquement évalué. Selon la méthode de lecture et menchu ou selon Kerber. Le titre est exprimé en doses de 50%. C'est ED50. Pour cette méthode de titrage, tout modèle biologique peut être utilisé, mais ce modèle doit être sensible au virus titré (cultures cellulaires, embryons, animaux de laboratoire). Selon l'effet infectieux des modèles biologiques infectés, ils sont répartis comme suit : cliniquement reconnus ; selon les changements pathomorphologiques ; au décès du modèle ; par l'accumulation d'hémagglutinine. Les résultats du travail dépendent de la dose du virus. Il a été établi que la dose du virus qui provoque 50 pour cent de l’effet infectieux est la moins sensible aux fluctuations et la plus déterminable de toutes les doses possibles. Le titre est exprimé en doses efficaces de 50 pour cent. Il s’agit d’une DE de 50. En fonction du modèle biologique utilisé et de l’effet obtenu, la dose de 50 pour cent peut être exprimée dans les unités suivantes : DL 50 – Cela représente 50 pour cent de la dose mortelle reçue par le laboratoire, qui est vivant en termes d'effet mortel. ID 50– il s’agit d’une dose infectieuse de 50 pour cent déterminée pour que le laboratoire soit vivant sur la base de signes cliniques ou de changements pathomorphologiques. DCE 50– il s’agit d’une dose létale embryonnaire de 50 pour cent déterminée sur les embryons de poulet par année de résultat. AID 50– il s’agit d’une dose infectieuse embryonnaire de 50 pour cent déterminée dans les embryons de poulet par des changements pathomorphologiques et l’accumulation d’hémagglutinine. TsPD 50– il s’agit d’une dose cytopathogène de 50 pour cent déterminée dans des cultures cellulaires par CPD. Si dans les systèmes infectés nous n'observons pas 50 pour cent de l'effet ID 50, alors le titre est calculé par lecture et menu : lg LD 50 = lg ECD - (% années d'ECD - 50%) / (% années d'ECD - % années de DPE) TOUT CELA MULTIPLIÉ PAR le facteur de dilution lg. Méthode 2 : sur l'effet infectieux du virus avec une évaluation de l'effet unique. Avec la méthode de formation de plaques en culture cellulaire, il y a un seul effet. Exprimé en unités formant variole ou en unités formant plaques PFU. Préparez une dilution de 10 fois du virus ; sélectionner un modèle biologique sensible ; Dans chaque dilution du virus, au moins 4 embryons sont infectés. Le titre est calculé à l'aide de la formule T=a divisé par V*n. a - le nombre moyen de pustules ou de plaques. V est le volume du contenu matériel = 0,2. n est le degré de dilution du virus. Méthode 3 : selon l'effet hémagglutinant du virus dans le GAEN. Ils ont mis RGA.
38.Principes du diagnostic en laboratoire des infections virales.
Les études de laboratoire jouent un rôle important dans l'établissement du diagnostic des maladies infectieuses, la prescription d'un traitement étiotrope et le contrôle de l'efficacité du traitement. Le processus de diagnostic spécifique en laboratoire repose sur l'identification de l'agent pathogène et de la réponse du corps humain au cours du processus infectieux. Elle comprend trois étapes : la collecte du matériel, son transport (épi n°39) et son étude en laboratoire : 1) La méthode virologique comprend deux étapes principales : l'isolement des virus et leur identification. Pour isoler les virus, des cultures cellulaires, des embryons de poulet et parfois des animaux de laboratoire sont utilisés. La présence du virus dans les cultures infectées est déterminée par le développement d'une dégénérescence cellulaire spécifique, c'est-à-dire effet cytopathogène, détection d'inclusions intracellulaires, ainsi que basé sur la détection d'un antigène spécifique par immunofluorescence, réactions d'hémadsorption et d'hémagglutination positives. Les virus sont identifiés par des méthodes immunologiques : réaction d'inhibition de l'hémagglutination, fixation du complément, neutralisation, précipitation sur gel, immunofluorescence. 2) Réactions sérologiques ; 3) Méthode immunologique (essais biologiques) ; 4) ELISA et PCR. Après réception des résultats de l'examen et prise en compte des données épidémiologiques et cliniques, un diagnostic définitif est établi.
39. Prise, préparation et transmission du brevet. Matériel pour virologue. Recherche.
Collecte, transport et examen de Pat. le matériel est réglementé par la législation vétérinaire. Lors de la prise, le tropisme du virus est pris en compte - la localisation préférée du virus dans une maladie donnée et la pathogenèse. Il est temps de se retrouver dans une impasse. matériel jusqu'à la fin de sa recherche - 2-4 heures. Si vous avez besoin de plus de temps, conservez-le (méthodes chimiques - solution à 50% de glycérol, physique - congélation), mais pas pour les luministes. microscopie. Transport vers spécial conteneurs avec accompagnement document et courrier. La préparation consiste à extraire le virus des cellules. Liquide matériau – filtration et centrifugation. Pour nettoyer les bactéries - bactériennes. filtres et antibiotiques (500-2000 unités pour 1 ml), pour champignons - fongicides (25 unités pour 1 ml), conserver 30-40 minutes, inoculer avec du pitata. environnement (aérobies – MPA, MPB, MPZh, anaérobies – Kitta-Tarozzi, champignons – Chapeka, Saburo). Matériau breveté dense : 1) prendre 1 à 1,5 g de matériau breveté ; 2) hacher avec des ciseaux ; 3) laver avec stérile. du verre ou du sable dans un mortier ; 4) suspension à 10 % avec une solution de Hanks ; 5) congeler et décongeler 2 fois ; 6)filtration à travers un filtre à gaze ; 7) centrifugation (3 000 tr/min, 15 min) ; 8) liquide surnageant - matériel contenant des virus, il est testé pour les bactéries (milieux nutritifs), des antibiotiques et des fongicides sont ajoutés.
40. Méthode microscopique de recherche en virologie.
1. Microscopie optique : 1) pour la détection du virus de la variole (méthode d'argenture de Morozov) ; 2) Détecter les corps d’inclusion (il s’agit d’une accumulation de virions ou de parties ou produits de la réaction de la cellule au virus ; ils peuvent être intranucléaires et cytoplasmiques) ; 3) détection du virus CPD (arrondi, fragmentation, mort) ; 4) détection des symplastes ; 5) travailler avec C/C ; 6) évaluation par sérologue. réactions (ELISA, RGAd, RTGAd). 2. Microscopie à luminescence : l'essentiel est que lorsqu'ils sont irradiés par des rayons UV, les atomes sont excités, puis passent à l'état initial avec libération d'énergie sous forme de rayonnement lumineux, son intensité s'apprécie en croix (vert émeraude = ++ ++ ; vert = ++ + ; vert-jaune = ++ ; jaune = + ; pas de lueur = –). Avant irradiation, la préparation est peinte avec des fluorochromes (FITC, acrédine orange, jaune, rhodamine). Il s'agit d'un simple fluorochrome placage. Complexe - MFA L'essence de la MFA est spécifique. interaction de l'anticorps avec le sérum marqué au fluorochrome (conjugué). 3.Microscopie électronique : 1) détection de tout virus ; 2) étude de sa taille, sa forme, sa structure, son type de symétrie, sa reproduction ; 3) étude de l'interaction du virus avec la cellule.
Au laboratoire de virologie, des travaux sont menés pour isoler les souches virales, leur identification et leur culture, et diverses études scientifiques sont réalisées. Lorsque vous travaillez avec des virus, vous devez tout d'abord :
1. Prévenir la contamination des souches virales par une microflore étrangère ;
2. Assurer la sécurité du personnel travaillant contre une éventuelle infection par des virus ;
3. Assurer la sécurité de la population environnante contre l'infection par des infections virales provenant des eaux usées, des cadavres d'animaux de laboratoire, etc.
Lors de l'étude de matériaux obtenus auprès de patients atteints d'infections virales, aux fins du diagnostic en laboratoire de ces maladies, diverses méthodes sont utilisées :
· Méthodes de microscopie électronique et, dans une moindre mesure, de microscopie optique ;
· Méthodes d'isolement et de culture de virus dans des cultures cellulaires ;
· Méthodes d'isolement et de culture de virus dans des embryons de poulet en développement et dans le corps d'animaux de laboratoire sensibles ;
· Identification des virus par leur capacité hémagglutinante ;
· Diverses méthodes de recherche sérologique : méthodes traditionnelles et expresses ;
· Méthodes de recherche en génétique moléculaire - hybridation moléculaire et réaction en chaîne par polymérase.
1.1.2. Matériaux étudiés pour les infections virales
Lors du prélèvement de matériel infectieux sur des personnes et des animaux, il est nécessaire de prendre en compte le tropisme des virus pour certains tissus et organes, la voie de libération du virus dans l'environnement extérieur et les caractéristiques de la pathogenèse d'une infection virale particulière.
Il existe des virus pneumotropes, entérotropes, hépatotropes, lymphotropes, neurotropes et dermotropes. Selon le tropisme, divers matériaux font l'objet de recherches. Par exemple, ils examinent le mucus de la gorge, les crachats, etc., si le virus est pneumotrope ; selles - avec des virus entérotropes; liquide provenant de vésicules ou de pustules, de croûtes - si le virus est dermotrope, etc.
1.1.3. Traitement du matériel contenant des virus
Les matériels infectieux, compte tenu du tropisme des virus et dans le respect de l'asepsie, sont placés dans des récipients stériles, soigneusement fermés et envoyés au laboratoire, placés dans un thermos avec de la glace.
Il est recommandé d'examiner le matériel le plus tôt possible, car les virus sont rapidement inactivés. La conservation du virus est facilitée en plaçant le matériel de test (dans une solution de glycérine à 50 %) au réfrigérateur à une température ne dépassant pas 5 o C. Mais la méthode la plus fiable est le stockage congelé à une température de -45 o C et moins. ; dans de telles conditions, le virus peut rester viable pendant longtemps.
Le traitement d'un matériau dense contenant des virus commence par son broyage dans un mortier ou par son broyage dans des appareils spéciaux - des homogénéisateurs. Ensuite, une suspension à 10 % est préparée dans une solution saline, qui est centrifugée à 2 000-3 000 tr/min pendant 15 à 30 minutes pour sédimenter les grosses particules. Les virus restent dans le surnageant, qui fait l’objet d’une étude plus approfondie.
Le matériel liquide contenant le virus est directement centrifugé et le surnageant est également obtenu.
En cas de doute sur la stérilité bactériologique du surnageant contenant le virus testé, des antibiotiques y sont ajoutés pour détruire les micro-organismes étrangers. Les antibiotiques n’affectent pas les virus et restent viables.
1.1.4.Méthodes de recherche microscopique en virologie
- Microscopie électronique
Les préparations électroscopiques sont préparées à partir de suspensions purifiées et concentrées contenant des virus ou de coupes ultrafines de tissus infectés par des virus. Les objets viraux sont appliqués sur des films substrats spéciaux placés sur des mailles de support. Les films de substrat doivent être très fins (pas plus de 30 nm d'épaisseur), transparents et suffisamment résistants, par exemple du carbone colloïdal. Les films sont appliqués sur des treillis de support en cuivre (2-3 mm de diamètre) percés de nombreux trous. Les médicaments sont ensuite traités de différentes manières.
Méthodes de pulvérisation de métal utilisé pour obtenir des agents de contraste. Les vapeurs de métaux lourds (or, platine, uranium, etc.), formées dans un appareil spécial sous vide et à haute température, sont dirigées sous un angle aigu vers le médicament contenant le virus. Les virus sont recouverts d’une fine couche de métal.
Méthode de contraste négatif est basé sur le fait que lorsque le médicament est traité avec certains sels de métaux lourds, par exemple une solution à 1-2% d'acide phosphotungstique, une couche plus dense est créée qui ne laisse pas passer les électrons et dans laquelle plus d'électrons -les objets transparents étudiés sont clairement visibles.
Coupe ultra fine combinée à un contraste négatif est le meilleur pour étudier la structure fine des virions et étudier les étapes d'interaction des virus avec la cellule, mais en même temps c'est le plus complexe. Les morceaux examinés de tissus infectés ou d'autres matériaux contenant des virus sont fixés dans un fixateur spécial (par exemple de l'osmium). Déshydrater par placement séquentiel dans des alcools de force croissante. Les échantillons sont remplis de plastique spécial, après polymérisation duquel se forment des blocs transparents solides. Des coupes ultrafines d'une épaisseur de 10 à 20 nm sont préparées à partir des blocs sur un microtome spécial. Les coupes obtenues sont contrastées en les plaçant dans une solution d'acide phosphotungstique.
Les préparations préparées par les méthodes décrites ci-dessus sont étudiées au microscope électronique à transmission dont la résolution atteint 0,2-0,3 nm. L'image de la préparation est observée sur l'écran fluorescent d'un microscope électronique et des plaques photographiques spéciales sont photographiées à partir desquelles des impressions sont obtenues. Grossissements reçus : ×100000-×400000.
La microscopie électronique à balayage est réalisée à l'aide d'un microscope électronique à balayage, dans lequel un mince faisceau d'électrons se déplace rapidement à travers l'objet étudié, c'est-à-dire scanne sa surface. En conséquence, un rayonnement d'électrons secondaires apparaît qui, passant à travers un tube cathodique, est converti en une image tridimensionnelle d'un objet sur un écran fluorescent.
La microscopie à balayage permet d'obtenir une image tridimensionnelle des virions (la préparation est d'abord pulvérisée de métaux), de distinguer les détails de la structure de leur surface, mais ne révèle pas leur structure interne. La résolution d'un microscope à balayage est de 7 à 20 nm.
- Microscopie optique
Au microscope optique, vous pouvez voir de gros virus dont la taille se situe dans la résolution du microscope - au moins 0,2 micron. Ainsi que des inclusions intracellulaires dans les tissus touchés par le virus.
Les gros virus, par exemple les poxvirus, et les inclusions sont détectés à l'aide de méthodes de coloration spéciales, en contraste de phase, dans un champ de vision sombre ; La microscopie fluorescente est également utilisée.
Les gros virus sont identifiés par coloration de Morozov (argenture). Pour identifier les inclusions intracellulaires, des coupes histologiques de tissus affectés, des frottis ou des empreintes sont préparés. Habituellement, les préparations sont colorées selon Romanovsky-Giemsa, parfois par d'autres méthodes. La détection des inclusions de Babes-Negri dans les cellules nerveuses du cerveau au cours de la rage revêt la plus grande importance pratique. A cet effet, les préparations sont colorées selon Mann.
Microscopie à luminescence. Les préparations préparées à partir de matériaux contenant de gros virus, des inclusions intracellulaires et des accumulations d'antigènes viraux sont colorées avec des solutions de colorants fluorochromes. En microscopie fluorescente sous lumière UV, les accumulations de virus génomiques à ARN colorées à l'orange d'acridine et les inclusions qu'elles forment sont visibles sous forme de granules rouges lumineux sur fond de cytoplasme cellulaire vert pâle ; Les virus génomiques à ADN dégagent une lueur vert émeraude.
Méthode d'immunofluorescence est basé sur la combinaison de virus, d'inclusions intracellulaires et d'accumulations d'antigènes viraux avec des anticorps antiviraux spécifiques marqués avec des colorants fluorochromes. Les complexes résultants brillent en microscopie à fluorescence.
 Portefeuille Qiwi - compte de réserve
Portefeuille Qiwi - compte de réserve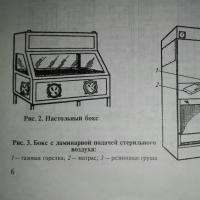 Méthodes de tests de laboratoire pour identifier les virus et caractéristiques de déchiffrement des résultats de la recherche
Méthodes de tests de laboratoire pour identifier les virus et caractéristiques de déchiffrement des résultats de la recherche Quel forum est le meilleur vBulletin ou PunBB
Quel forum est le meilleur vBulletin ou PunBB Premiers pas dans l'optimisation des moteurs de recherche Forum Simple Machines : suppression des droits d'auteur et des liens externes Sujet php de l'index cru alimenté par smf
Premiers pas dans l'optimisation des moteurs de recherche Forum Simple Machines : suppression des droits d'auteur et des liens externes Sujet php de l'index cru alimenté par smf Mods Gta 5 comment installer un personnage
Mods Gta 5 comment installer un personnage fichiers Télécharger enregistrer pour gta 5
fichiers Télécharger enregistrer pour gta 5 Assistance technique Apple russe – expérience de communication personnelle
Assistance technique Apple russe – expérience de communication personnelle